| 6. Változékonyság | TARTALOM | I. alosztály: Egyhüvelyűek (Monodelphia) |
EMLŐSÖK
FEJEZETEK
Az emlősök éppúgy a gerincesek állattörzsébe tartoznak, mint a madarak, csúszómászók, kétéltűek és halak. Éppen azért, mielőtt az emlősök részletes tárgyalásába fognánk, néhány szóval meg kell emlékeznünk a gerinces állatokról is.
A gerinceseket (vertebrata) a hossztengely irányában két egyenlő – jobb és bal – részre lehet széthasítani, úgyhogy egyik rész tükörképe a másiknak; éppen azért kétoldali részarányos állatoknak nevezzük őket. Természetesen a kétoldali részarányosság nem mindig tökéletes; gondoljunk csak az emlősök kissé baloldalra tolódott szivére, vagy a jobboldalon fekvő májra és az össze-vissza gombolyodott belekre. A kétoldali részarányos félhez hasonlóan, egy hát és hasi – de már nem egyenlő – félre is tagolható a gerinces test, melyek közül mindig a háti részben találjuk a központi idegrendszert, a hasi vagy zsigericső pedig a tengőéleti szerveket foglalja magában. Rátekintésre, külsőleg a legtöbb gerinces állaton három tájat különböztethetünk meg, nevezetesen a fej-, törzs- és farktájat; ez utóbbi sok gerincesen elmaradhat. A törzshöz kapcsolódnak a páros elrendezkedésű végtagok, melyek hiányozhatnak is, de négynél több számuk nem lehet. A gerincesek bőrét az irha szemölcsös része jellemzi, mely szemölcsök a pikkelyek, tollak, szőrök képzésében játszanak nagy szerepet. A gerincesek legjellemzőbb tulajdonsága azonban, hogy belső, szilárd, porcos vagy csontos tengelyük van, amely a gerincesek főzöménél egyes részekre csigolyákra (vertebrae) tagolt. A gerincoszlop tagoltsága a gerinces test szelvényezettségével függ össze, melyet nemcsak a csigolyák, hanem az ér, ideg- és izomrendszer is – többé-kevésbbé – mutatnak. A vér teljesen zárt edényhálózatban kering és a vér piros színét a benne úszkáló sejtek színe szabja meg. A lélegzőszerv – legyen az kopoltyú vagy tüdő – a szájüreggel áll szoros összefüggésben.
Az emlősöket éppúgy jellemzi a szőr, mint a madarakat a toll. Minthogy melegvérű állatok, a test melegének megóvása szempontjából védőburokra van szükségük. A védőburok a bőrből és függelékeiből áll, melyeket együttesen köztakarónak nevezünk.
A köztakaró mint zárt burok, elsősorban védőberendezés és hőszabályozó, ezenkívül mint a lélegzőszerv, el-, illetve kiválasztószerv is bír jelentőséggel, a mellett, hogy a tapintás érzékét is szolgálja. A bőr három rétegből áll: a hám (epidermis), irha (corium) és bőralatti kötőszövetből (subcutis). Ezek közül a középső, az irha, vastagságánál fogva a legfeltünőbb. Amint a közönséges életben bőrnek nevezünk, nem egyéb, mint az emlősállat irhája, mely a cserzés révén tartóssá és szívóssá lett. A hámréteg alsó nedves rétegét, nyálkás réteg néven különböztetjük meg a hám felületes részétől, mely utóbbinak mélyebben fekvő sejtjei cseppfolyós, felületesebben fekvő sejtjei pedig megszilárdult szaruanyagot tartalmaznak; a legkülső réteg korpa vagy pikkely alakjában válik le a test felületéről. Ahol az elhalt sejtek leválását a sűrű szőrzet akadályozza, ott a korpaképződés mindig feltűnő. A bőr színe a hám legmélyebb rétegeiben és az irhában levő sötétbarna színű festékszemcsék (pigment) mennyiségétől függ. A bőralatti kötőszövet sok zsírszövetet foglal magában, amely egyrészt a test melegének szigetelésére, másrészt a táplálék tartalékolására szolgál.

A hámréteg erős utánpótlást kívánó legkülső rétege mindig újból képződik és pedig a mélyebb nyálkás rétegből, melynek legalsó részét az irhából belényomuló szemölcsök hálózatosan széttagolják és amely réteget felfedezőjének tiszteletére Malpighi-féle recézetnek (rete Malpighii) nevezték el; a hám táplálását az egyes irhaszemölcsökbe alulról behatoló véredények végzik.
A bőr függeléke a szőr. A szőr alsó vége, vagyis a gyökere, a bőr mélyedésébe, az úgynevezett szőrtüszőbe illeszkedik be, mely mélyen az irhába húzódik; a szőrtüsző fenekén a szőrszemölcs emelkedik ki, amelyre süvegszerűen illeszkedik rá a szőr gyökerének legalsó megvastagodott része, a hagyma. A szőrnek a bőrből kinyúló része, a szőr nyele, hámsejtekből áll, melyek közül a belső lágyabbak a velőt, a külső elszarusodottak a kérget adják; a velő gömbölyded és levegőtartalmú, a kéreg rostalakúan meghosszabbodott sejtekből áll; a gyapjúszőrben nincs velő. A szőr színe, mely a legvilágosabb fehértől a legmélyebb feketéig mindenféle árnyalatú lehet, festékanyag és levegőtartalmától függ; ősz szőrökben csak levegő van.
A szőrtüszők a testnek nagy területén ugyanabban az irányban ferdén türemkednek be, s így a szőrtüszőkből kiálló szőrök is, szabályosan házfedélcserépszerűen borulnak egymásra, ami által sokkal jobban őrzik meg a test melegét. A szőr bőrbe süllyesztett szárán egy akaratunktól független izom – a hajmeresztő – tapad meg; hogyha az izom félelem, ijedtség vagy izgatottság következtében összehúzódik, fölmereszti a szőrt, bőrünk lúdbőrös lesz és hajunk az égnek áll. A szőrdűlés iránya a testen nem egyforma; általában felülről lefelé és elülről hátrafelé, a testtartásnak és a mozgás irányának megfelelően dűl. A lajhár a legjobb példa arra, hogy mennyire befolyásolja a testtartás és mozgás a szőrdűlés irányát, mely egész nap fejjel és háttal lefelé, hasával az égnek fordulva csüng a fán, mely állandó testhelyzetnek megfelelően a szőrök a hastól a hát felé dűlnek; természetes, hogy ez az egészen különös szőrdűlés a lajhár életmódja mellett a legcélszerűbb.
Kidd W. szerint a szőrdűlés az állatok passzív és aktív szokásaihoz alkalmazkodik, vagyis a test állásának, a test és testrészek mozgásának és a törzs és végtagok egymáshoz való viszonyának függvénye. Így keletkeznek a különféle szőrmezők és szőráramlatok, amelyeknek találkozási helyén, tarajok, szőrfogók jönnek létre. Ez utóbbiak keletkezésének okai azonban nem mindig szembetűnőek.
Még érdekesebb, miképpen rendeződnek az egyes szőrök a bőrön csoportokba. A csoportosulás jelensége helyesen értelmezve, igen érdekes adatokat szolgáltat a szőröknek a pikkelyekhez tollakhoz és egyéb bőrképződményekhez való viszonyról. A kérdéssel Weber Miksa amszterdami zoológus foglalkozott behatóbban, akinek az emlősökről írott könyve, más emlőstani kérdésekben is igen gyakran használt és nélkülözhetetlen alapvető munka. Weber a pikkelyekből indul ki, azokból a lapos, házfedélcserépszerűen egymásra fektetett szaruképződményekből, melyeket a hasonló módon elrendezett irhaszemölcsök választanak ki és amelyek a külső erőművi behatások ellen védelmezik az állat testét. A reptíliák szarupikkelyeit (kígyóbőr) jól ismerjük és tudjuk, hogy a tollak sem egyebek hővédő szervekké átalakult pikkelyeknél, melyek a madárlábon mint pikkelyek maradtak meg. Az emlősök körében valóságos szarupikkelyeket a tobzoskáknál találunk; csakhogy míg a kígyó kinőtt szarupikkelyeit levedli és az új pikkely gyorsan pótlódik, addig az emlősöknél ilyfajta vedlés nem következik be, az emlős szarupikkelye az elhasználódás fokához és az állat növekedéséhez mérten, fokozatosan gyarapodik.
Az öves állatok hámrétegének elszarusodása, az irharéteg elcsontosodása mellett, kisebb mértékű; a szarupikkelyek között és mögött szőrcsoportokat találunk. Weber szerint tehát pikkely és szőrözet egymást nem helyettesítik, ahol pikkely van, ott szőrzet is lehetséges, nem úgy, mint például a madárlábon, ahol vagy toll, vagy pikkely van jelen. És bár a szőrök keletkezése a hüllők szarupikkelyeire vezethető vissza, pikkely és szőr egymás mellett is fejlődhetnek. A törzsfejlődés folyamán a szőr a pikkelyeket valószínűleg a szükségesség mértékében szorította ki, minél több hővédelemre és minél kevesebb mechanikai védelemre volt az állatnak szüksége, annál több szőr és kevesebb pikkely keletkezett. Weber, de Meyere tanulmányai alapján, rámutat arra, hogy némely emlősön, amelynek rég nincsenek pikkelyei, a szőrök elrendezése ma is olyan, mintha az állat pikkelyes volna. Ilyen „pikkelyesen” elrendezett a szőrzet az erszényesek, rágcsálók, rovarevők végtagjain és farkán; különösen feltűnő a foghíjasoknál, ahová a tobzoskák is tartoznak. A sörényes hangyász dúsan és zászlósan szőrözött farka, még nagy, fekete szarupikkelyekkel födött.
Az emlős szőrcsoportok, mintha pikkelysorok között volnának, rendszerint az állatnak és végtagjainak hossztengelyére keresztben állanak; legtöbbször három szőr alkot egy csoportot, ezek közül egy erősebb – középső szőr –, kettő gyengébb – oldalszőr. Vedlés után, ezek mellett, a szőrtüszők kiöblösödése útján, mellékszőrök keletkezhetnek, amely mellékszőrök a gyapjúszőrnek, míg az eredeti szőrök a fedőszőröknek felel meg. A sűrű és lágy gyapjúszőr, a mérsékelt és hideg égöv alatt élő emlősök téli bundáját jellemzi; tavasszal a fedőszőrrel együtt, a gyapjúszőr kis kihull és a gyapjúszőr csak ősszel nő ki újra. A szőrruha cseréje tehát évenként kétszer történik meg, helyesebben, az emlős minden évben kétszer vedlik. Régebben azt hitték, hogy csak tavaszi vedlés van, és hogy például a havasi nyúl téli fehér bundája a színes nyári öltözet kifakulása útján keletkezik; ma már tudjuk, hogy ősszel is van vedlés. Vedlés előtt egyedül a baribal bundája fakul feltűnően fahéjszínbarnára. Hasonló, bár kisebb mértékű színváltozást vedlés előtt házinyulaink bundáján is észlelhetünk, mely színváltozás a szőröknek vedléselőtti csökkentett táplálkozásával áll kapcsolatban. Ennek megfelelő az az eset is, hogy vedlés előtt némely feketeszínű madár tolla rozsdásszínű lesz. Csak tisztán tropikus és magas északi állatok érik be talán évenként egyszeri, fokozatos és lassú vedléssel; így az állatkertek antilopházaiban sohase találunk oly tömegesen kihullott szőröket, mint a szarvas- és bölénykarámokban a tavaszi vedlés alkalmával; ezenkívül minden szűcs tudja, hogy a jegesmedve bundája télen, nyáron, tavasszal, ősszel egyformán „jó”; hasonlóképpen a víziemlősök gereznája bármely időben „használható”. A speciális célokat szolgáló tapintó- vagy sinus-szőrökről – amelyek szőrtüszőinek kötőszöveti rétegében vérrel telt öblök, sinusok, találhatók –, később, a tapintással kapcsolatban lesz szó.
A különösen vastag és merev disznóserte, átmenet a sündisznó és tarajos sül tüskéihez, melyek rendszerint szőrrel keverten fordulnak elő; ez esetben tehát a külső mechanikai behatások megakadályozására a szőrök alakultak át védő szervekké. Nagyon természetesen a tüskék is, keletkezésük és szerkezetüknek megfelelően, különfélék lehetnek.
Az emlős testén még más bőrképződmények is vannak. Némely helyén a testnek a hámréteg – amely amúgy is hajlamos a szarusodásra – rendkívül megvastagodott és elszarusodott. Ilyen szaruképződmények: a csőrös emlős „csőre”, a teve mellén levő szarúkérgek, a ló szarú gesztenyéi, az oroszlán farktüskéi a hangyászsün szarúsarkantyúi; még a rinoceroszok tekintélyes orrszarva is ide sorolandó, mely nem egyéb, mint egy rostos szerkezetű szarutömeg. A szaruképződményeket az alatta levő irha – mely ezeken a helyeken mirigyeket rendszerint nem tartalmaz – erőteljesen kifejlődött, vérdús, szemölcsös rétege választja ki.
Más módon keletkeznek az antilopok, kecskék, juhok, szarvasmarhák tülkei és a szarvasok agancsai. A szarvak képzésében az irhának egy bőrcsontja játszik nagy szerepet; ebből a bőrcsontból másodlagosan fejlődik az évente váltódó szarvasagancs; az egész agancsot, mellékágaival együtt, kezdetben szőrös bőr – a háncs – vonja be, mely az agancs teljes kinövése után lassan elszárad és a csontos szárakról lefoszlik. Az üres szarvúak szarva, tülke, annyira lényeges szerepet játszik az állat életében, hogy állandóan a fejen marad; a tülök a homlokcsont szarvnyúlványán, a szarvcsapon ül, a szarvcsap irharétege választja ki; lényegileg tehát nem egyéb, mint egy tömött csontcsapra ráhúzott, üres szaruképződmény, amelynek alapján viselőit (antilop, kecske, juh, szarvasmarha) Cavicornia – üres szarvúak – néven foglaljuk össze. Ellentétüknél a szarvasoknál – Cervicornia – az agancs megfelel az üres szarvúak szarvcsapjának, a mulandó szőrös bőr – a háncs – a tülöknek. Bizonyos mértékben az agancs és az üres szarv között helyezkednek el a villásszarvú antilop és zsiráf szarvai; nevezetesen a villásszarvú antilop szarva bár üres tülkű: elágazó, sőt tülkét évenként leveti az állat; a zsiráf szőrrel bevont szarva legjobban a szarvas agancshoz hasonlít, csontcsapja mégis állandóan a helyén marad, sőt a csontcsapot borító szőrös bőr is állandó.
Általában véve, úgy látszik, hogy az agancs levetése nem a szarvasfélék kizárólagos sajátsága. Az üres-szarvúaknál is (borjú, antilop, vadjuh) megfigyelték, hogy az első, fiatalkori tülök leválik; hasonlóképpen az állatkertekben tartott indiai orrszarvún észlelték, hogy az orron levő szarv minden négy-öt esztendőben leesik és újra képződik.
A szárazföldi emlősök egész életében rendkívül fontos szerepük van, a végtagok végén található szaruképződményeknek, amelyek csak a víziemlősöknél (fókák és rokonaik) szorúltak háttérbe, és csak a ceteknél hiányzanak. E szaruképződmények alapján az összes szárazföldi emlősök két nagy csoportra oszthatók, a szerint, hogy az ujjak végét, köröm és karom, vagy pata és csülök fedi; a végtagok használata és az emlősök mozgása is szorosan összefügg az újjak végén lévő szaruképletek alakjával és minőségével. Legkevésbbé födi és védi az utolsó ujjpercet a köröm (félmajmok, majmok, ember), amelyet az előre húzódott és megnagyobbodott ujjpárna – a tapintásra való ujjhegy – az ujj végének hátsó felére (dorzalis oldalára) szorított vissza.

A ló patás ujjának hosszmetszete. Queisser rajza után.
A patás állatokat kivéve, valamennyi szárazföldi emlős karmokat visel. A karom hosszirányban erősen megnyult; erősen ívelt és kétoldalról összehajló lapja az alsó lágyabb, könnyen lekopó szarutalpot keskeny szalaggá szorítja össze. Közvetlenül a karom mögött van az ujjpárna; járás közben az ujjon járó emlősöknél csak az ujjpárnák, a talponjáróknál az ujjpárna mellett a talpgumók is érintik a földet.
A köröm, karom és pata folyton nő, hogy az állandóan erős kopást ellensúlyozza. A köröm az irhának egy erősen szemölcsös részéből, a körömgyökérből vagy pártából nő ki, amely a körmöt körülvevő körömsánc alatt fekszik; a növekvő köröm, a körömágy fölött halad az ujjhegye felé. A körömsánc, mely tulajdonképpen a köröm megerősítésére való, legkevésbé a patán fejlődik ki, minthogy a pata, mint teljesen zárt szarutok, amúgy is elég szilárdan függ össze a bőrrel és a végső ujjperccel.
A karomperc – a karom különösen tartós megerősítése céljából –, hosszában hasadt is lehet, mely esetben a karom alapja léc alakjában a hasítékot is kitölti és úgy szilárdabban ül a kétfelé hasított karomperc között: ilyen a karmuk az ásó-életmódot folytató (vakondok, tobzoskák) állatoknak. A lajhár és hangyász karomperce azonban – ez állatok más módon használják erősen karmaikat – csak hosszában árkolt.
A patásállatok végső ujjperce lényegesen különbözik a többiétől, amennyiben erősen megvastagodott és kiszélesedett. A pata fali részlete, mely a köröm lapjának felel meg, a véső ujjpercet kétoldalról egészen körülveszi és éles szögben zárul a szarutalphoz; a közbeeső részben a nyír foglal helyet, amely egy elszarusodott ujjpárnának felel meg. A legtökéletesebben fejlett a ló patája; ezt szokták, mint a pata típusát is emlegetni; típusos patája van a ló és rokonai mellett a tapírnak és orrszarvúnak is; csak az elefánt és szirtiborz lábainak szaruképződményei egészen különlegesek és egyedül állók.
A bőrhöz tartoznak még a benne előforduló mirigyek is, amelyek a madárbőrrel szemben, az emlősbőrre igen jellemzők. A mirigyek eloszlása az egész bőr felületén egyenletes is lehet, de lehet olyan is hogy bizonyos helyeken a mirigyek felhalmozódnak és mirigy-szervekké alakulnak át. Így minden egyes szőrszál tüszőjével faggyúmirigyek állnak összeköttetésben; a hajmeresztőizom összehúzódásakor nyomást gyakorol a mirigyre, mely tartalmát kiüríti és bezsírozza a szőrt és a bőrnek körülötte fekvő részeit; a zsír kitűnően védi a bőrt az átnedvesedés ellen.
Az egyszerű mirigyek közül a verejtékmirigyek éppúgy jellemzik az emlősöket, mint a szőrtüsző faggyúmirigyei; a faggyúmirigyektől nemcsak a kiválasztott anyag minőségében, hanem a kiválasztás módjában és felépítésének mikéntjében is különbözik. A verejtékmirigyek a hőmérsékletet szabályozzák; ha a külső nagy meleg folytán izzadni kezdünk, verejtékmirigyeink bizonyos sós nedvet, izzadságot vagy verítéket választanak ki, mely a bőr felületén elpárolog s ezáltal lehűti a testet. A veríték vízből, sóból és illó zsírsavakból áll, mely utóbbi, gyors elpárolgás esetén, a nem mindig kellemes, de sokszor jellemző izzadság szagot okozza. Az izzadtság-mirigy egyszerű csöves mirigy, mely mélyebben, a bőralatti kötőszövetben foglal helyet, ahol az el nem ágazó mirigy csöve gomolyt alkot; kivezető csövük a hámrétegben dugóhúzószerűen csavarodott és a bőr felületén nyílik a szabadba.
A faggyúmirigyek a test bizonyos részein nagyon felhalmozódnak és hatalmas, igen erőteljesen működő mirigyzacskókat vagy csoportokat alkotnak. Sok ilyen mirigycsoport található az emlősökön, melyek legtöbbje a nemi szervek közelében helyezkedett el, jeléül annak, hogy a nemi élettel állnak szorosabb összefüggésben.
Legjellemzőbb mirigyei az emlősbőrnek a tejmirigyek; elsősorban a nőstényeket jellemzik, de csökevényesen a hímben is megtalálhatók. Általánosan ismert dolog, hogy a tejmirigyek kivételesen a hímeken is működésbe léphetnek; mezőgazdasági kiállításokon nem egyszer látható a svájci nemes Sanne kecskefajta, melynek hímjei is tejelnek. Különbséget kell tennünk a kloákás emlősök (csőrös emlős, hangyász sün) és a többi emlős tejmirigyei között, nemcsak azért, mert a kloákás emlősök testük sajátos felépítésénél fogva is egészen külön állók, hanem azért is, mert tejmirigyeik az izzadságmirigyekhez, a többi emlősöké a faggyúmirigyekhez hasonlóak. Bizonyára ez a különbség is hozzájárult ahhoz, hogy a kloákás emlősöket a többivel szembeállítsuk; azonban Bresslau E. legújabb vizsgálatai szerint („A csőrös emlősök, erszényesek és placentás emlősök szoptató szerve”, melynek első fejezete „Az Echidna szoptatószervének fejlődése és eredete” – 1907), a hangyászsün nőstény-magzatának tejmirigye, csak fejlődésének legkezdetén, hasonlít a körülötte levő izzadságmirigyekhez, amelyeket a tejmirigyek fejlődésükben feltűnően gyorsan túlszárnyalnak. Fontos feladatuknak megfelelően „rövidesen hatalmas mirigycsövekké alakulnak, amelyekben már világosan felismerhető a tejmirigy”. Vagyis csak a tejmirigy kezdeménye egyezik meg az erszény bőrének izzadság mirigyeivel, aztán céljuknak megfelelően másmódon fejlődnek tovább.


A kloákás emlősöktől eltekintve, amelyeken a tejmirigyek elszórva és egyenként torkollanak ki egy lapos bőrmezőre, minden más nőstényemlősön, a szoptatás tartamának idejére, a tejmirigyek megnagyobbodnak. A tejmirigy bimbója olyképpen keletkezik, hogy a mirigymező a körülötte levő bőrsánccal együtt megnyúlik, vagy pedig csak a mirigymező emelkedik ki a bőrsáncon túl; mind a két esetben a tejmirigyek kivezető nyílásai igen közel fekszenek egymáshoz, sőt egyesülhetnek is; az így keletkezett – többé-kevésbé golyóalakú szervet – emlőknek (mammae) nevezzük.
Az emlők fejlődése a magzat tejvonalára vagy tejlécére vezethető vissza. A tejvonal a magzaton az elülső végtagok kezdeményétől, a lágyék tájáig terjed. A tejlécen tejdombok keletkeznek s a köztük levő tejléc-részletek később felszívódnak. Ezekből a tejdombokból fejlődnek ki a tejmirigyek és az emlőbimbók, melyeknek száma és elhelyezése, az ivadékok számával, a táplálék felvételének módjával és az állat testtartásával függ össze. Az emlő száma 22–2 között váltakozik; elhelyezése szerint lehet mell, has és lágyéktáji tejmirigy, bár kivételesen a háton, hónaljban, sőt a combokon is elfordulhatnak.
Az emlők szerkezetével, fejlődésével és az egész tejmirigy-problémával Gegenbaur, a heidelbergi nagy anatómus foglalkozott először behatóan.
Gegenbaur megtanít először is arra, hogy a valódi és az álbimbó között különbség van; nevezetesen, ha a mirigymező kiemelkedik környezetéből és olyan bimbóvá alakul, melynek csúcsán nyílnak a mirigycsövek, akkor valódi bimbóval van dolgunk; ha azonban csak a mirigymezőt körülvevő bőrsánc emelkedik ki és alakul át bimbóvá és a bimbó kivezető csövének alján szájadzanak a tejmirigyek, akkor álbimbó keletkezik.
")
Gegenbaur az emlők szerkezetét az állítólagos emlőtáskával hozta összefüggésbe, amely szerv, a törzsfejlődés folyamán, mint a szaporodás segédszerve, látszólag igen nagy szerepet játszott; Gegenbaur elméletét tanítványa és munkatársa, Klaatsch, származástani kutatásai közben még ki is bővítette és kiegészítette. Bresslau legújabb vizsgálatai azonban egészen más eredményekre vezettek, mert megállapította, hogy a hangyász-sün szoptató szervei, keletkezésüket tekintve, semmiféle összefüggésben sincsenek az emlőtáskával és hogy „az emlőtáska fogalma végérvényesen törlendő”. Az emlőtáska fogalmának helyébe, Bresslau „elsőleges kezdeményei” lépnek, amelyek segítségével az emlősök származását is, sokkal könnyebben érthetjük meg. Ezek az elsőleges kezdemények „az embrionális élet egész korai szakaszában” lépnek fel és pedig „az egyébként teljesen egyforma hasbőr epidermiszén, két, hosszanti, lécalakú megvastagodás” keletkezik, amely a hossztengely irányában, a köldöknyílás két oldalán húzódik. Ezek „abban az időben, amikor a testfalak záródása a köldöktájon bekövetkezik, megakadályozzák a bőrizomzat kiterjedését, a hasbőr középső részére; így elsősorban okai annak, hogy bőrizommentes hasbőrterület keletkezik, amely a későbbi erszénynek felel meg”. Bresslau az elsőleges kezdeményeket – és ezeknek a hasfalléceknek ez az általános törzsfejlődéstani jelentősége – „költőszervek maradványainak” tartja, „melyek az emlősök (reptiliaszerű) őseinél, hasonló módon voltak kifejlődve, mint ma a madaraknál”. „Az emlősök tejkészülékei a gerincesek ezen legmagasabb csoportjának nem teljesen új szerzeményei, hanem olyan ősrégi állapotokkal vannak összefüggésben, amilyenek a tojástrakó, még nem emlős állatoknál az ivadékgondozás szolgálatában voltak kifejlődve. A tojásrakástól és költéstől, az eleven szülésig és a szoptatásig való átmeneti állapotok sajátságos átalakuláson mentek át, amely átalakulás tette lehetővé, hogy azok az új körülmények között is az ivadékgondozás szolgálatában maradjanak.”
A szaporodás és ivadékgondozás módja éppúgy jellemző az emlősökre, mint a többi állatokra. Az emlősök egy ideig szoptatják kicsinyeiket, vagyis a már ismertetett tejmirigyek folyékony váladékával, a tejjel táplálják azokat. A tej az újszülött továbbfejlődéséhez szükséges összes anyagokat tartalmazza. Az emlőből szoptatás igen jellemző az emlősökre. A madarak közül a tejhez hasonló étellel – a begytejjel – etetik kezdetben kicsinyeiket a galambok is, amely azonban a begy belső többrétegű hámjából levált, zsírcseppekkel telt sejtekből áll.
Az emlősök elevenszülők. A pete az anya testében fejlődik csírává és az a csíra magzattá. Az anya testében marad a magzat mindaddig, míg a külső élethez szükséges fejlettségi fokot el nem érte. Szüléskor a magzatburkok felrepednek, a magzat és burkai között az összeköttetés megszűnik, olyképpen, hogy a köldökzsinór elszakad, vagy az anyaállat elharapja azt; a magzatburkok az úgynevezett utószülés alkalmával hagyják el az anya testét. A csúszómászók, kétéltűek és halak között is vannak elevenszülők, melyek tulajdonképpen csak a tojásrakással késnek el, a tojás az anya testén belül kel ki és a kicsinyek így elevenen jönnek a világra; a legtöbb elevenszülő fajnál azonban a tojás tényleges lerakását mesterségesen elő lehet idézni. Mindezekben a kivételes esetekben azonban, az aránylag nagy tojás, bár az anya testén belül fejlődik, a fejlődéséhez szükséges anyagokat és erőket a már eleve benne levő alakító- és táplálószikből meríti.
Nem így az emlősöknél, ahol a pete és a magzat táplálása az anya testének anyagaiból a magzat burkok segítségével történik. Ezen tulajdonság alapján, mely az emlősök fejlődéstörténetére is rendkívül jellemző, az emlősöket megkülönböztethetjük az összes többi gerincesektől. Az ausztráliai kloákás emlősök azonban „tojásrakó emlősök”, mely elnevezés már önmagában is ellentmondásnak látszik. Azonban, mint ahogy a csúszómászók, kétéltűek és halaknál késleltetett tojásrakásnál, úgy ez esetben korai szülésről beszélhetünk, éppúgy, mint ahogy kétségtelenül koraszülők a kengurú és más erszényesek is; rég bebizonyított dolog, hogy a kloákások pergamenthéjba zárt petéje – „tojása” – a pergamenthéjon belül, a körülvevő héjjal együtt növekedik oly módon, hogy közben az anyai test nedveiből táplálkozik.
Az emlősállatok petéje mikroszkópikus kicsinységű képződmény, egyszerű sejt, mely egészen más megjelenésű, mint a madarak petéje, a tojás, mert sem szilárd héja, sem számottevő szíktartalma nincsen; így nem csoda, hogy jó időnek kellett eltelnie addig, míg az omne vivum ex ovo – minden élő tojásból lett – régi elvét, az emlősökre is érvényesnek találták.
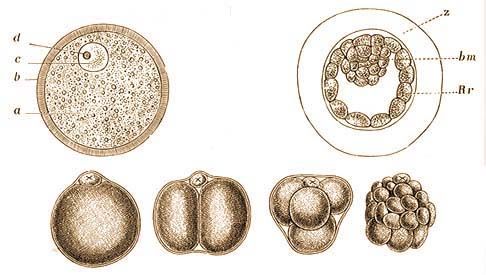
| A házinyúl kifejlett, de még nem egészen érett petéje. | A vakond hólyagalakú csírája hosszanti metszetben. |
| a a fénylőöv, b a pete szíkje, c a csírahólyag, d a csírafolt. | z fénylő öv, bm barázdálódási maradék, Rh Rauber-féle réteg. |
| Az egérpete osztódásának néhány jellemző szaka. Mihalkovics után. | |

Az egysejtű emlős petét – mely sajátos képzőerőt és fejlődésképességet tartalmaz – 1828-ban Baer Károly Ernő – nagynevű orosz természetbúvár fedezte fel egy nőstény kutya petevezetékében. Régebben az emlős állat petefészkében lévő petetömlőket, az úgynevezett Graaf-féle tüszőket tartották petéknek, noha csak e tüszők belsejében képződnek a peték. A véredényekkel gazdagon körülfont tüsző belsejében a pete bizonyos idő mulva megérik; ilyenkor a tüsző fala felreped és a pete a méh nyálkahártyájának vérzése kíséretében (sárlás, folyatás, menstruáció) kiszabadul.
A megtermékenyített pete további fejlődésében az a legcsodálatosabb, hogy egy erekben gazdag szíktömlő képződik, bár az emlőspete tápláló-szíket nem tartalmaz. A szíktömlő-kialakulását sokáig nem tudták megmagyarázni; ma már bizonyosra vehető, hogy az emlősök ősei szíkben gazdag és héjjal bíró tojásokkal szaporodtak, akárcsak a madarak és csúszómászók; a szíktömlő tehát, mint örökség maradt az emlősök őseitől a mai emlősökre, olyképpen, hogy a pete szíkje a törzsfejlődés folyamán megfogyott, a szíkhólyag fejlődési módja azonban megmaradt.
Az anya testén belül a fejlődés folyamán még más különös változások is történnek a magzaton; minden változás azonban megmagyarázható abból a szükségességből, hogy a szíkszegény csírát az anyának véréből kell táplálnia. Hogy ez megtörténhessen, úgy az anya mint a magzat testén nyálkahártyák képződnek, melyek nagy felületen és többé-kevésbbé bensőségesen érintkeznek egymással; e hártyafelületeken át, a test nedvei osmosis útján, kicserélődnek egymással; a magzat és az anya testét tehát emlősrendenként különféle és többé-kevésbé bonyolult berendezések kötik össze. Mint már említettük a „tojásrakó” emlősök tojása az anyai testen belül is növekedik; növekedés közben a tojás pergamentszerű héja kitágul és az anyai test belső nyálkahártyái által kiválasztott táplálónedvet átereszti.
A magzat táplálásának legfontosabb szerve – a madaraknál és csúszómászóknál is meglevő – és az összes többi emlősökre jellemző hugytömlő (allantois); csakhogy míg a madaraknál és csúszómászóknál csak a gázcserét bonyolítja le, addig az emlősöknél a gázcserén kívül az anyagcsere forgalmát is el kell látnia. A húgytömlő a magzat hátsó részéből véredényekkel együtt ered s bőven áterezett; a magzat védelmére szolgáló magzating (amnion) és a legkülső irhahártya (chorion) között terjed el, az említett két hártya közötti részt többé-kevésbbé kitölti; a húgytömlő nyele, a benne levő véredénytörzsekkel együtt, a köldökzsinór főtömegét alkotja

[A tojásban fejlődő állatoknak nincs chorionjuk: achoria (madarak, reptiliák, kloákás emlősök és erszényesek). A legalacsonyabbrendű gerincesekben (halak, kétéltűek) az amnion is hiányzik; anamnia, mert teljesen a méhen kívül fejlődnek; ezek az ovipara. A tojástrakó állatok petéi a méhben termékenyíttetnek meg és már fejlődésnek indulnak, amikor lerakják őket: ovovivipara. Az eleven szülők a fejlettebb embriókat hozzák a világra: vivipara.].
A legegyszerűbb és legtökéletesebb módja a magzat táplálásának, hogy a külső magzatburok felületén szétszórtan apró bolyhok keletkeznek, melyek segítségével a fejlődő magzat az anyaméh (uterus) nyálkahártyájához tapad. A legtöbb erszényes magzata ezen a fokon marad s a magzat burkai és az anyaméhe között sohase létesül bensőbb összeköttetés. Ilyen körülmények között a magzat táplálása nagyon tökéletlen, s minthogy fejlődése csak egy bizonyos fokig haladhat előre, a terhesség ideje rövid s a kicsinyek igen fejletlen állapotban születnek meg. A magasabbrendű emlősök magzatburkainak felülete a méhhel a méhlepény (placenta) útján függ össze; ennek egyik része a méh nyálkahártyájának sarjadzása útján keletkezik – a méhlepény anyai része –, másik része a külső magzatburok felületéről nyúlik be – a méhlepény magzati része.

Az összeköttetést a külső magzatburok (chorion) bolyhai létesítik, melyek a méh nyálkahártyájának megfelelő mélyedéseibe illeszkednek bele. Az összeköttetés a méh és magzat között nagyon különböző és az egyes emlős rendekre jellemző. A méh falának nyálkahártyája és a magzatburkok össze is nőhetnek egymással; ez esetben a szülés vérzéssel jár, hiszen a magzatburkokkal együtt a méh fala is eltávozik. Ezeket hullóburkosoknak (deciduata) szokás nevezni, szemben a hullóburkot nélkülözőkkel (adeciduata), amelyeknél a különböző hártyák összeköttetése nem ilyen bensőséges.

A tehén párnás méhlepénye. m anyaméh, bb bolyhos burok, p az anyaméh méhlepénypárnája, p' a magzat méhlepénypárnája. Hertwig O. után.

Szülés előtt a magzat vérkeringése egészen más, mint szülés után, mert míg előtte az anya vérkeringésével van szoros összefüggésben, utána, helyesebben a tüdők működésbe lépése után, az újszülött egyéni vérkeringése lép működésbe. A magzat fejlődéséből jegyezzünk meg annyit, hogy fejlődés közben átmegy az úgynevezett hal-állapoton, mikor a páros kopoltyúíveknek megfelelően párosan elágazó fővéredényei vannak és amikor a szív éppúgy csak egy egyszerű kamrából áll, mint a halaknál; átmegy a reptilia-állapoton, amikor szívkamrái éppoly tökéletlenül vannak egymástól elválasztva, mint a krokodilusnál, és éppen ezért a magzat reptilia-állapotában, a vénás és artériás vér keveredik; vagyis az emlős-magzat egyéni fejlődésében átmegy mindazokon a fokozatokon, amelyen maga a törzs is fejlődés folyamán már általment, és amely fokozatok a gerincesek egyes csoportjait jellemzik.
Az emlősök és madarak melegvérű állatok, helyesebben állandó testhőmérsékletű gerincesek, testük elég magas hőmérséklete tehát a külső környezet, a levegő hőfokától független és állandó; ezzel szemben a hidegvérű, helyesebben változó testhőmérsékletű állatok (csúszómászók, kétéltűek és halak) hőfoka változó és a külső környezet hőmérsékletével van szoros összefüggésben. A külső környezettől független testhőmérséklet csak tökéletes vérkeringés és megfelelő hővédő-berendezések mellett lehetséges. Tökéletes vérkeringés mellett gyorsabb az anyagcsere, gyorsabb a tápláló anyagok „égésének” folyamata, fokozottabb a test „fűtése”; kitűnő hővédő-berendezés a madarak tollazata és az emlősök szőrözete.
A vérkeringés akkor tökéletes, ha a szénsavat és más bomlástermékeket tartalmazó vénás vér, és az oxigénben és tápanyagban dús artériás vér nem keveredhetnek egymással. Keveredés pedig nem lehetséges, mert úgy a két szívpitvar, mint a két kamra válaszfallal tökéletesen el van választva egymástól. Az úgynevezett nagy vérkörben a vérkeringés a bal kamrából indul ki, ahonnan az artériás vér eljut a test minden részébe; a hajszálerekben megtörténik az anyag- és gázcsere és az elhasznált – vénás – vér a gyüjtőerek útján bejut a jobb pitvarba s innen a jobb kamrába. Az úgynevezett kis vérkörben a jobb kamrából a tüdőbe jut a vénás vér, ahol a hajszálerekben megtörténik a gázcsere – a szénsav eltávozik, helyébe oxigén lép – innen az artériás vér eljut a bal pitvarba s a bal pitvarból kiindulási helyére, a bal kamrába. Érdekes, hogy a kis vérkör mozgását szabályozó jobb kamra és bal pitvar kevésbbé izmosak, mint a nagy vérkört szabályozó szívrészeké, ami érthető, hiszen csak a közvetlen közelben lévő tüdőbe kell a vért eljuttatniok, a nagy vérkörben mozgó vérnek pedig a test legtávolabbi részeit is át kell járnia. A szív olyképpen működik, hogy előbb egyidejűleg a két pitvar és azután egyidejűleg a két kamra húzódik össze; a kamrákból a vérnek a pitvarokba való visszajutását a szívbillentyűk akadályozzák meg.
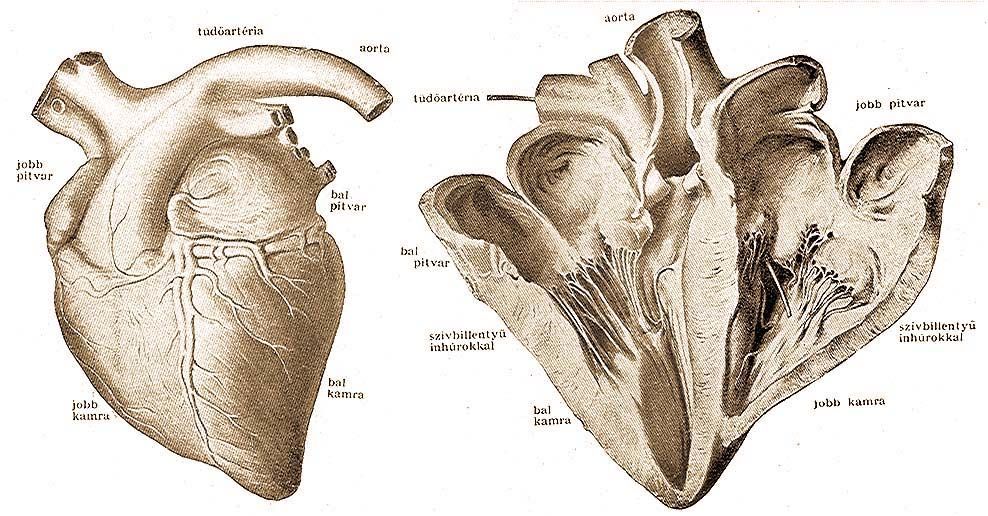
| A szív képe a nagy vértörzsekkel. Roloff rajza után. | A szív belseje, a kamra, pitvar, nagy vértörzsek, szívbillentyűk és ínhúrokkal. Roloff rajza után. |
A gyors anyagcsere, a gyors égés s így az erősebb fűtés a lélekzéssel, helyesebben az oxigén-felvétellel áll kapcsolatban. Az önmagában színtelen vérfolyadékban – vérplazmában – igen nagyszámú, mikroszkópikus kicsinységű, kerekded, piros színű sejt – ezek okozzák a vér piros színét – úszkál, amelyeket vörös vértesteknek nevezünk. A vörös vérsejt rendkívül fontos anyagot tartalmaz, a haemoglobint, mely az oxigén és széndioxid tulajdonképpeni tovaszállítója. A haemoglobin ugyanis rendkívül könnyen köti meg az oxigént és elszállítva azt a szövetekhez, leadja, hogy az ott keletkezett bomlás terméket, a szénoxydot felvehesse, amelyet viszont a tüdőben bocsát szabadon. A gázcsere érintkezési és felületi hatáson alapszik. Könnyű belátni, hogy minél több vérsejt úszkál a vérben, annál nagyobb az érintkezési felület és ezzel arányosan több oxigént vehet fel a vér. Egy köbcentiméter emberi vérben körülbelül 4 1/2 millió vörös vérsejt van, míg ugyanannyi békavérben csak fél milliónyi vörös vérsejtet találunk; kétségtelen azonban, hogy a béka vörös vérsejtjei sokkal nagyobbak, mint az emberéi.
Említettük, hogy a melegvérűség az emlőst és madarat egyformán jellemzi; a madarak vére azonban még melegebb, mint az emlősöké, mert anyagcseréjük is gyorsabb; mind a két tulajdonság valójában a madarak rendkívüli mozgékonyságával függ össze, amelyet viszont jóval kisebb termetük tesz lehetővé. A madár és emlős vérkeringésében egy alaktani különbséget is találunk; nevezetesen a fejlődés folyamán úgy a madárnál, mint az emlősnél meglévő páros kopoltyúívekből, helyesebben az ennek megfelelően haladó fővérerekből, a madaraknál egy jobboldali ág, az emlősöknél egy baloldali ág maradt meg, mint a szívből kiinduló főér (aorta). Minthogy az emlősök főere baloldali, valószínű, hogy a baloldali agyfélteke jobban táplált; minthogy a baloldali agyféltekében van a jobboldal mozgató-idegeinek központja, az emberek általános jobbkezűségét is ezzel hozzák kapcsolatba. A melegvérűség egyetlen hátránya, hogy a melegvérű lény élete a test melegének meglehetősen szűk korlátai közé van szorítva. Nagyon kevés emlős állat alszik téli álmot, mert a téli álom merevségét nagyon kevés emlős bírja ki; hiszen az aránylag oly kisméretű hőemelkedés, amilyen láz esetén jelentkezik, már életét veszélyezteti. Gondoljunk csak a lázhőmérőre és arra, hogy ezzel kapcsolatban az ember élete is a hőmérsékletnek milyen szűk határai közé van szorítva.
A lélekzéssel és anyagcserével függ össze és csak az emlősöket jellemzi a mell és hasüreget elválasztó rekeszizom. A rekeszizom a mellüreg nemes szerveit – tüdőt, szívet –, a hasüreg belső szerveitől tökéletesen elválasztja; a mellüreg felé boltozatszerűen domborodó falát mindössze csak a nyelőcső, néhány ideg- és véredény fúrja át; izomzata a szélek felől, a középpont felé sugarasan elrendezett. A rekeszizom a belélekzés legfontosabb izma, összehúzódáskor lelapul, a mellüreg tehát tágul, szívóhatást fejt ki; kilélekzéskor ellazul, miáltal ismét a mellüreg felé domborodik; görcsös összehúzódása a csuklás; köhögés és tüsszentésnél szintén közreműködik, működésének megszünte halálra vezet. A lélekzés tulajdonképpen a tüdőben történik, amely a külvilággal, a szájon és orrnyíláson át, a gégecső és hörgők útján van összeköttetésben; a légvétel a mellüreg tágulásával és szűkülésével történik, tehát abban – többé-kevésbbé – a mellkast alkotó bordák és ezek mozgató szervei is részt vesznek.

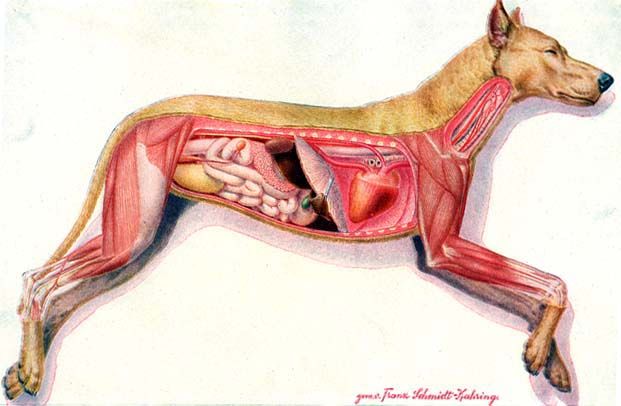
Egy nőstény kutya belső szervei
A nyirok és nyirokedényrendszer éppoly fontos szerepet játszik az emlősök életében, mint a vér és a véredényrendszer, bár általában véve, csak betegségek és oltás alkalmával szoktunk róla tudomást venni. A nép „testnedvnek” nevezi, a valóságban igen fontos tápláló és kiegyenlítő folyadék. A vese megbetegedésekor a kiválasztás elégtelen volta miatt túlságosan sok nyirok halmozódik fel testünkben s ilyenkor vízibetegségről beszélünk; öröklött „rossz” nyirokösszetételnél skrofulózis keletkezik, és ha betegségokozó paraziták behatolása által „elromlott”, akkor vérmérgezés következik be. A nyirok a vérhez hasonló szerkezetű, plazmájában túlnyomóan színtelen vérsejtek vannak; az elhalt nyiroksejtek alkotják a gennyet. A nyirok nagy jelentősége tehát kétségtelen. A nyirok tartalmazza az anyagcsere termékeit is; a bélbolyhok falán, hámján, a bélben folyékony állapotban lévő tápanyagok – zsírok, fehérjék és sók – áthatolnak, felszívódnak és végeredményben a bolyhok tengelyében haladó nyirokérbe és onnan a bélcsatornát felfüggesztő bélfodor (mesentherium) nagyszámú nyirokcsomóiba jutnak; a tápláló-anyagokkal telt testnedv fehéres és tejszerű; a neve bélnyirok, (chylus).
Nyirokcsomók helyett nyirokmirigyekről is szokás beszélni, bár a legtöbb nyirokcsomó nem mirigy, mert nem választ ki semmit. Nyirokmirigynek tekinthető a lép. A lép vérképző szerv, amelyen a szervezet egész vérmennyisége, aránylag rövid idő alatt megy keresztül; benne a vörös vérsejtek elpusztulnak és színtelen, fehér vérsejtek (leukocyta) fejlődnek, melyek amoeba alakúak és amoeboid mozgást végeznek; tulajdonképpen a gerinctelen állatok vérsejtjeinek felelnek meg. Feladatuk rendkívül fontos és különös, nevezetesen a testbe hatoló betegségokozókat – baktériumokat – semmisítik meg. Mihelyt a baktériumok seben keresztül, vagy a fertőzésnek más útján a testbe jutottak, a fertőzés helyéhez legközelebb álló nyirokcsomó megduzzad – így például „elmérgesedett” ujjsebnél a könyökben és a hónaljban lévő – és ez az állapot addig tart, míg a fehér vérsejteknek sikerült a kezdetben szörnyű gyorsasággal szaporodó baktériumok összességét megenni.
A nyirokmirigyekhez, helyesebben a vérképző szervekhez sorolható a magzat- vagy csecsemő-mirigy (thymus) is; a szív és mellcsont között foglal helyet, csecsemő korban tekintélyes képződmény, a felnőtt állaton azonban rendesen visszafejlődik. Leghatalmasabban fejlett talán a borjúban, ahol 400–600 gramm súlyú is lehet; konyhai műnyelven egyszerűen borjúmirigynek – németül Briese, franciául ris de veau – nevezik.
Itt említjük meg a paizsmirigyet (thyreoidea) is, amely félholdalakúan elől a nyakon, a gégefő alatt fekszik. Tulajdonképpeni feladata még ismeretlen, csak veszedelmes hatásait ismerjük. Így a közismert golyva nem egyéb, mint betegesen elfajult és megnagyobbodott paizsmirigy. A golyvás ember puffadt külsejű, dülledt szemű és rendesen gyöngeelméjű is; a golyvát operációval nem lehet, helyesebben nem szabad tökéletesen kiirtani, mert teljes kiirtása, még súlyosabb következményekkel jár, mint a betegség és olyan működésbeli zavarok léphetnek fel, melyek a beteg halálát okozhatják. Az így fellépő működésbeli zavarok, arra engednek következtetni, hogy a pajzsmirigy – minthogy hozzá erős véredények tapadnak – az agyvérnyomás szabályozására szolgálhat, avagy pedig arra, hogy a testben keletkezett úgynevezett anyagcseremérgeket távol tartsa a fejtől. Ez utóbbi esetben működése a nyirokmirigyekéhez volna hasonló.
Végeredményben a testnek valamennyi nyirokere két főtörzsbe ömlik. A mellvezeték (ductus thoracicus) a hátulsó végtagok, a has és a medence falának és zsigereinek, a mellkas, az elülső bal végtag, a fej és a nyak balfelének, nyirokereit szedi össze és az elülső üres véna baloldali részébe torkolik. A másik, a jobboldali nyiroktörzs (truncus lymphaticus dexter) a fej és a nyak jobbfeléből, továbbá az elülső jobb végtagból gyűjti össze a nyirokereket és az elülső üres vénába jobboldalt ömlik. A nyirokedény rendszernek tehát külön mozgató szerve, nyirokszerve nincsen, hanem közvetlenül a véredényrendszerrel függ össze s így közvetlenül és állandóan ömleszti tartalmát, a nyirkot a vérbe.
A csontváznak hármas feladata van, nevezetesen szilárd vázat alkot, a test üregeinek képzéséhez járul hozzá s így védi az azokba foglalt szerveket, és végül az izmok, ínak tapadására is szolgál. Ezeken kívül egész sereg olyan tulajdonsága is van, mely az emlősök mozgásának módjával és az élelemszerzés mikéntjével függnek össze.



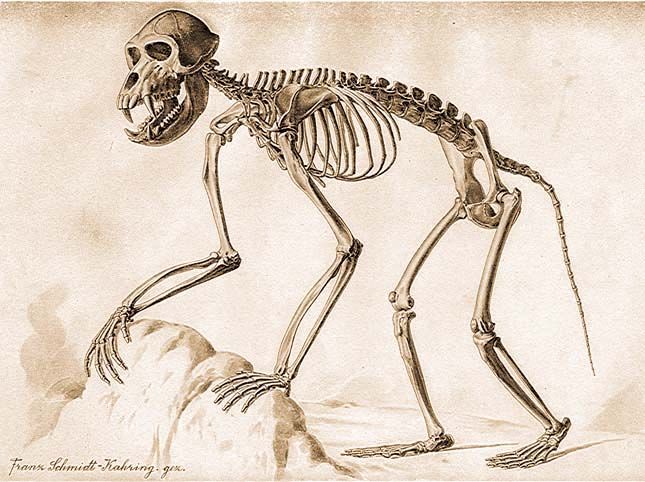
Egy pávián csontváza
Az emlős koponya nyakszirt csontja (occiput) két bütyökkel ízesül az első nyakcsigolyához, a fejgyámhoz (atlas). A koponya kettős bütyke az összes gerinces állatok között csak a kétéltűeken ismétlődik s így közelfekvőnek látszik az a gondolat, hogy a kétéltűek és az emlősök között valamilyen származástani összefüggés lehetséges. A koponya ízesülésének ez a sajátos módja, függőleges irányban a fejnek csak bólintó mozgását engedi meg; a fej vízszintes irányú mozgásait a fejgyám és a második nyakcsigolya, a fejforgató (epistropheus) sajátságosan átalakult ízülete teszi lehetővé. A madarak feje egy bütyökkel izül a fejgyámhoz, ezen kívül a fejgyám és fejforgató közti izülete is olyan, mint az emlősöké; ez a magyarázata annak, hogy a madarak fejüket tetszésszerinti irányba tudják forgatni; hozzájárul ehhez az a sajátság is, hogy a madarak nyaka hosszú, nyakcsigolyáik száma 8–23 között váltakozik. Ezzel szemben az emlősök nyakcsigolyáinak száma állandóan hét; még a zsiráfnak sincs több, pedig elég hosszú a nyaka; a hetes szám még a ceteknél is kimutatható, pedig a ceteknek nincs külsőleg felismerhető és mozgatható nyakuk és nyakcsigolyáik ennek megfelelően többé-kevésbbé megrövidültek, sőt össze is nőttek. Kivétel ez alól csupán a lajhárnak egyetlen egy neme, amelynél a nyakcsigolyák száma 10 is lehet; ez esetben a nyak abszolút értelemben vett megnyúlása, kétségtelenül az állandóan kúszva lógó életmóddal függ össze, mely életmóddal kapcsolatban az állat fejének szabadabb mozgásra van szüksége.

A szentbernáthegyi kutya első és második nyakcsigolyája. A forgócsigolya fognyujtványa a fejgyám nyilásba jól látható.
Más gerinces állattal összehasonlítva, az emlősön nyomban felötlik, hogy mind a két pár végtagját meglehetősen egyformán használja, csupán azokon a fajokon alakultak ki az elülső végtagok különlegesen és csenevészedtek el a hátulsók, amelyek a levegőben való mozgáshoz, vagy a vízi élethez alkalmazkodtak. A két végtagpár alkata azonban korántsem oly elütő, mint a madaraknál, hol mindegyik pár más-más közegben való mozgás feltételeinek felel meg. Épp ezért az emlősök lába, a szárazföldi gerincesek ötujjú alaptervének megfelelő és ha attól eltér (egy patások, két patások), akkor a változás mind a két végtagpárt egyaránt és ugyanabban az irányban érinti.
Itt néhány szót kell szólanunk a szembe helyezhető hüvelykujjal bíró kézről, arról a kézről, amelyről az elfogulatlan laikusnak az az érzése, hogy ezáltal lesz az ember második teremtővé, mert ez teszi őt képessé arra, hogy a természetet alkotóan utánozza. És ha meggondoljuk, mennyi mindenre használható az emberi kéz és milyen csodálatos a teljesítőképessége az értelem irányítása és finom beidegzése következtében, akkor ez igaz is. És mégis, hogy a kéz törzsfejlődéstanilag nagyon régi képződmény – mondja Vogt Károly – már abból is kitűnik, hogy a kézen is az ősi ötujjas állapot maradt meg. Ahogyan mi ezt az ősi ötujjas állapotot, az alacsonyrendű – hidegvérű – gerinceseknél ismerjük, úgy az csak a származástanilag is régi és alacsonyrendű erszényeseknél tér vissza, ezeken kívül pedig csak a megint nagyon régi és legtöbbnyire csak az ősrégi madagaszkári szárazulatra szorítkozó félmajmoknál, és végül a majmoknál található. Csakhogy ezeknél a kéz már megváltozott – tovább fejlődött – amennyiben a hüvelykujj visszafejlődött, amit világosan láthatunk abból, hogy nagysága kisebb, állása megváltozott s ehhez képest teljesítménye sem egyenlő egy jól fejlett hüvelykujj teljesítményével; ebből a már visszafejlődött állapotból az emberi kéz nem alakulhatott ki, már csak azért sem, mert a visszafelé haladó fejlődés lehetetlen. Másrészt az ember, a röpdösés kivételével, mindent tud, amit az emlősök tudnak; tud szaladni, ugrani, kúszni, úszni, ásni; de mindent rosszabbul tud, mint éppen azok az emlősök, amelyek a fenti tulajdonságok egyikét egyoldalúan bár, de egyirányú fejlődésük következtében nagyszerűen tudják; az embernek ezt a kezdetleges fokon álló sokoldalúságát sokan az ember származásával hozzák összefüggésbe. Klaatsch és mások legújabb származástani kutatásaik alapján arra az eredményre jutottak, hogy az ember eredetét sokkal mélyebben kell keresni az emlősök törzsfáján, mint eddig tették.
A hátulsó végtag végső része a láb is azt bizonyítja, hogy az ember és legközelebbi állati rokonai is, alapjában véve sok tekintetben a testi fejlődésnek egy ősrégi fokán maradtak meg. A már említett Vogt Károly ezt származástanilag a következőképpen magyarázza: Az ember, a majmok és félmajmok, továbbá a medvék és a legalacsonyabb rendű emlősállatok: az erszényesek és kloákások, talponjárók, azaz járás közben talpukra lépnek, vagyis sarkuk is érinti a földet. Ezzel szemben az emlősök legnagyobb része ujjonjáró, azaz lábtőcsontjuk meghosszabbodott, sarkukat járás közben – többé-kevésbé – a levegőben hordják és csak ujjaikkal érintik a földet. Vogt megemlíti még, hogy az emlősök legrégibb kihalt képviselői, akármilyen csoporthoz tartoztak, mint azt megkövesedett maradványaik bizonyítják, talponjárók voltak. Tehát nemcsak az öt ujjú kéz – mondja Vogt – hanem a talponjáró láb is egészen kezdetleges ősi bélyeg. És lényegileg mégis csak ez a láb teszi lehetővé azt, hogy az ember egyenesen jár, melynek következtében feje másképpen fejlődött és kezei szabadokká váltak.

A gerincoszlopon különösen az ágyékcsigolyák nagy mozgékonysága tűnik szembe. A földön és vízben való mozgás sokfélesége s különösen a törzsnek változatos hajladozása, az ágyék könnyű mozgathatását kívánja meg, holott a madarak vázán a törzs merev egészet alkot s így mozgásuk nehézkesebb.
Az elülső végtagokat a gerincoszlophoz függesztő vállöv az emlősnél a szabadabb mozgásnak megfelelően egészen más alkotású, mint más gerinces állatokon. A vállöv elülső részén ugyanis csak a kulcscsontpár fejlődött ki és a hollóorrcsont, csak a lapockacsont elülső végén lévő rövid hollóorrnyujtvány (processus coracoideus) képében ötlik fel. A vállövnek még messzebbmenő elsatnyulását tapasztaljuk azokon az emlősökön, melyek elülső végtagjaikat éppúgy használják, mint a hátulsókat (ugrásra, futásra). Ebben az esetben a kulcscsont vagy teljesen hiányzik, vagy pedig rugalmas, középen kissé megcsontosodott szalaggá lett. Ezeknél a test ruganyosan lóg a mellső lábuk között, ami futás közben és hirtelen megállás esetén, rendkívül előnyös. A mellső végtagot oldal felé is mozgatható emlősökön (ásók, repülők), a kulcscsont is hatalmasan fejlett. A vállöv legnagyobb csontja általában mindig a lapocka, melyet még erőteljesebbé tesz a hosszában haladó tövis (spina scapulae) és az erősen fejlett vállcsúcs (acromium) is.
A mellkas a madarakéhoz képest fejletlenebb. Az emlősök szegycsontjai kicsiny és a hozzácsatlakozó bordák alsó fele porcos, holott a madaraké nemcsak a mellett, hanem a has egy részét is beborítja és a bordák megcsontosodott alsó fele, úgy a bordával, mint a szegycsonttal mozgékonyan egyesült.
A fejváz és a fogazat alkata a legszorosabban összefügg a táplálék felvételének módjával. Az emlősök az egyedüli gerincesek, melyek táplálékukat, mielőtt lenyelnék, gondosan szétdarabolják és megrágják. A préda megragadása és felaprózása, megrágása csak úgy lehetséges, hogy az arckoponya szilárd összefüggésben áll az agykoponyával; az arcváz szilárdságát fokozza, hogy a mozgatható alsó állkapocs közvetlenül illeszkedik a koponyához s így csupa szilárd részre támaszkodva, erőteljesebben fejtheti ki hatását. Csak az agykoponya egyes csontjai olvadnak későn össze; ez azonban szilárdságát nem csökkenti, mert az egyes koponyacsontok, egymásba dugott két fésű módjára, zegzugosan, úgynevezett varratokkal (sutura) még mindig elég szilárdan csatlakoznak egymáshoz.

A fejváz csontjainak ez a késői összenövése, az agyvelő lassú, és sokáig tartó növekedésével kapcsolatos – ismeretes, hogy az összes gerincesek között az emlősöknek van a legfejlettebb agyvelejük – mert az agyvelő növekedését az agytoknak is követnie kell.
Az alsó állkapocs eredetileg két – egy jobb és egy bal – darabból áll, melyek a legtöbb emlősnél – a rágcsálókat kivéve – elől igen erősen függnek össze egymással; az alsó állkapocs hátsó felén, egy-egy erősebb nyúlvány van, amelynek segítségével ízülnek a koponyához. Azok a csontok: a négyszögcsont (os quadratum) és az ízületi csont (os articulare), amelyek az alsóbbrendű gerinceseken az alsó állkapcsot lazán függesztik a koponyához, az emlősökön hiányozni látszanak. Ha azonban az emlőscsíra fejlődését figyelemmel kísérjük, látni fogjuk, hogy ezek a csontok is megvannak, csakhogy a fejlődés folyamán a fülbe húzódtak s ott hallócsontokká módosultak. És valóban! Mert míg az összes többi gerinceseknek – a halakat kivéve – csak egy hallócsontjuk van, addig az emlősöknek három, amelyeket alakjuk után: ülő-, kalapács- és kengyelnek nevezzük; a hallócsontok továbbítják a dobhártyától a hangot a tulajdonképpeni belső fülbe. A kétéltűeknél, csúszómászóknál és madaraknál is meglévő egyetlen hallócsontocskának a kengyel, míg az ülőnek és kalapácsnak, a négyszögcsont és izületi csont felel meg.

Ez az eset példa arra, miképpen gazdálkodik a természet az öröklött anyaggal, miképpen változtatja át az öröklött tulajdonságokat, a szükségnek megfelelően; más szóval az alkalmazkodás kényszerűsége oly hatalmas erő, mely az öröklött tulajdonságokat is megváltoztatja.
A fogazat a test minden szerve között a legnagyobb szívóssággal öröklődik át. Még azoknak az emlősöknek is, amelyeknek a születés után kivételesen nincsenek fogaik (kloákások, néhány foghijas és cetek), születés előtt fogkezdeményeik vannak, amelyek ez esetben rövidesen eltűnnek, felszívódnak. A kihalt madarak között fogazottak is vannak, melyeket épp ezért fogasmadaraknak (Odontornithes) neveznek; az alsóbbrendű gerincesek fogai számban ingadozók, az állkapcson kívül a szájüreg más csontjain is (szájpadlás, kopoltyúív) nagyobb számban jelenhetnek meg és alakban, valamint szerkezetben sem oly tökéletesek; némely foghíjas fogazata – nem minden foghíjas „fogszegény” is – annyira egyszerű és kezdetleges, hogy egészen az alacsonyrendű gerincesek fogazatára emlékeztet, jeléül annak, hogy a fejlődésben nagyon visszamaradt.

Majom koponyája fogváltás közben. Roloff rajza után.
A legtöbb emlős fogazatában a munkamegosztás elve érvényesül: a különféle fogcsoportok különféle munkát végeznek, melynek megfelelően alakjuk is módosul, erősen vagy gyöngén fejlődnek ki, vagy teljesen hiányozhatnak is, a szerint, hogy a táplálék szerzésben milyen feladat – sok, kevés vagy semmilyen – jut az egyes csoportoknak. Igen sok és teljesen egyforma kúpalakú foga van – mint amilyen a csúszómászóké szokott lenni – az emlősök közül a delfinnek, melynek alsó fogai a felsők közé, a felsők viszont az alsók közé „vágnak”. Ez a fogazat csak a zsákmány megragadására alkalmas; mihelyt a zsákmányt fel is kell aprítani, a munkamegosztás elvének megfelelően, különféle fogakat találunk.
Az emlősök fogazatában alak- és elhelyeződés szerint általában véve háromféle fogat különböztetünk meg.
A metszőfogak (incisivi) az állkapocs elején foglalnak helyet és elsősorban a táplálék megfogására, leharapására és csak részben a feldarabolására is szolgálnak. Alakjuk többé-kevésbé lapos és vízszintes éllel bíró, de lehetnek más alakúak is, mint például az elefánt agyara. Közös megállapodás alapján, metszőfogaknak nevezik azokat a fogakat, amelyek az állközti csontba vannak beékelve; ez a meghatározás igen jó, csakhogy szószerint csak a felső metszőfogakra vonatkoztatható, mert az alsó állkapocsfelek között nincs állközti csont, úgyhogy ez esetben főképpen alakjuk és helyzetük az irányadó.
A szemfogak (canini) a metszőfogak mögött foglalnak helyet, az állkapocsnak ama részében, ahol az hátrafelé fordul; ennek megfelelően, minden állkapocsfélben csak egy-egy szemfog van, amely rendszerint erőteljesebb is. A szemfog igen gyakran veszedelmes „fegyver” a ragadozóknál és más harcias állatoknál is.
A zápfogak az állkapocs hátulsó, szemfog mögötti részén ékelődnek be. Feladatuk a táplálék felaprítása, megrágása, megőrlése; feladatuknak megfelelően alakjuk is különböző. Hátrafelé rendesen nagyobbodók, meglehetősen összetettek: már korántsem oly egyszerűek, mint a szem- és metszőfogak; így a húsevők zápfoga tarajos, a rovarevőké hegyescsúcsú, a mindenevőké gumós, a növényevőké pedig többé-kevésbbé lapos. A zápfogak közül az elsőket – rendesen egyszerűbbek is – előzápfogaknak (praemolares), a hátsókat valódi zápfogaknak (molares) nevezzük.
Ezzel kapcsolatban eljutunk a tejfogakhoz és a fogváltáshoz. Nevezetesen előzápfogaknak azokat a zápfogakat nevezzük, amelyek már a tejfogazatban is megvoltak és valódi zápfogaknak azokat, amelyeknek tejfogelőzőik nem voltak. A tejfogazat tehát mindig kevesebb fogból áll, mint az állandó végleges fogazat. És ez nagyon természetes is, hiszen időközben a csecsemő felnőtté serdült és kis állkapcsai is megfelelő nagyságra növekedtek; s minthogy a fogak növekedése korlátozott és így nem tart lépést az állkapocs növekedésével, a megnövekedett állkapocsban szükségszerűen több fognak kell lennie. Az emlősök tejfogazata és fogváltása alapjában véve csak átmenet abból az ősi állapotból, amilyent az alacsonyabbrendű gerinceseken még ma is találunk. Nevezetesen ezeknél ahányszor kitörnek vagy elkopnak a fogak, annyiszor fejlődnek újra s így megtörténhetik, hogy egy állkapocsban teljesen különféle fognemzedék van egyszerre. Nem így az emlősöknél, fogcsere csak egyszer történhet s az is csak bizonyos időben és rendesen meghatározott sorrendben következik be. Természetesen a szabály alól vannak kivételek is, ilyen kivételesen és nagyon sajátságosan történik a fogváltás például az elefántnál.
A fognak az állkapocs medrébe szilárdan beékelődött részét foggyökérnek nevezzük, melytől könnyed befűződés – a fog nyaka – által válik el a szabadon kiálló fog korona. A fog eredetét tekintve, nem egyéb, mint a bőrnek egy bőrcsontosodása, amennyiben az írhának egy szemölcsén keletkezik, mely süvegszerűen választja ki a csonthoz hasonló fogállományt (dentin), amely a fog belső ürege körül párhuzamos csövecskék alakjában rakódik le.
Vannak azonban olyan fogak is, így például a rágcsálók metszőfogai, sőt némelyik rágcsálónak a zápfogai is, amelyeknek szemölcse állandóan működik, s így növekedésük határtalan lenne, ha az állandó kopás nem szabályozná nagyságukat. Azonban a legtöbb emlősnek a foga, bizonyos nagyságot elérve, annyira lefűződik a szemölcsről, hogy az összeköttetés éppen csak a táplálás fenntartására elegendő, de a fog növekedése be van fejezve; a kifejlett foghúsból kiemelkedő részét, mely a rágásnál oly nagy szerepet játszik, a koronát, rendszerint kemény, rideg anyag, a fogzománc borítja; a foggyökérben nincs zománc, minthogy az csak a fognak az állcsontban való megrögzítésére szolgál.
Az emlős-fogazat alakja függ az öröklött tulajdonságoktól, de függ az alkalmazkodás körülményeitől is, úgyhogy a fogazat végeredményben e két egymás ellen működő alaperő eredőjeként alakul ki. Általánosságban feltehetjük – és nagyon sok bizonyíték is támogatja ezt a feltevést – hogy a tejfogazat inkább az öröklött tulajdonságokat mutatja, míg az állandó fogazat, a felnőtt állat életmódjához alkalmazkodott. Az emlős-fogazatból tehát éppoly joggal vonhatunk következtetést a származásra és a rokonságra, mint az állat táplálékára és egész életmódjára. Ezek szerint az emlős rendszertanban – és pedig az élőknél éppúgy, mint a kihaltaknál, mely utóbbiakból igen sok esetben csak a fogakat ismerjük – a fogazatnak rendkívül nagy szerepet kell játszania.
Könnyebb áttekinthetőség kedvéért a fogazatot fogképletben szokás kifejezni és pedig tört alakjában; a törtnek számlálója a felső fogakat, nevezője pedig az alsókat jelenti; a sorrend a természetesnek megfelelő, vagyis első helyre írjuk a metszőfogakat, második helyre a szemfogakat és harmadik helyre a zápfogakat; ez utóbbiakat két számmal is jelölhetjük, mely esetben külön írjuk az előzápfogakat és külön a zápfogakat. A törtek elé a fogak latin nevének rövidítéseit is (i, c, pm, m) ki szokás tenni.
Az anyagcsere szervei közül a táplálkozás műszereit kell szemügyre vennünk. A táplálék megragadásában, a fogakon kívül, az emlősök osztályára többé-kevésbbé jellemző mozgékony ajkak is részt vesznek; működésüket kivételesen az orr (elefánt) vagy a nyelv is (hangyász) pótolhatja.
A mag- és húsevő madarak begyéhez hasonló, táplálékot megpuhító, mirigyes előcsarnokot az emlősöknél nem találunk. Némely majmoknak és rágcsálóknak pofazsebeik vannak, amelyek azonban csak a táplálék elraktározására, ideiglenes megőrzésére valók. Az emlős állat, az emésztés bevezetéséül, fogaival megrágja, megőrli táplálékát és e közben a szájüreg mirigyeinek váladékával keveri össze, úgyhogy a táplálék megemésztése már a szájüregben indul meg. A váladékot a szájüreg számtalan kis mirigyén kívül, melyek a nyálkahártyán mindenütt előfordulhatnak, három pár nyálmirigy szolgáltatja.
A megrágott tápláléknak a nyállal való összekeveredést egy fölötte izmos és mozgékony szerv: a nyelv végzi. Minthogy a nyelv rendszerint fölötte hosszú is, kinyujtható a szájból s így igen alkalmas arra, hogy az állat azzal saját és kicsinyeinek testét tisztogassa; néha a táplálék megfogására is (zsiráf) szolgál. A nyelv azonban érzékszerv is; a nyelv hegyén és testén van a szemölcsös rész, sokszor elszarúsodott szemölcsökkel (macskák érdes nyelve); gyökerét a tüszős rész borítja, amelyen az úgynevezett körülárkolt és leveles szemölcsök (papillae circumvallatae és foliatae) találhatók; e kétféle szemölcs mélyedéseiben vannak az ízlelő kelyhek vagy ízlelő bimbók.

Ízlelőbimbók, erősen nagyítva. Queisser rajza után.
Az orr- és szájüreget a vízszintes kemény szájpadlás választja el egymástól, amelynek hátrafelé egyenes folytatása a lágy szájpadlás vagy ínyvitorla, mely a szájüreget a garatüregtől (pharynx) színházi függönyhöz hasonlóan választja el; alsó-szabad szélének közepén emelkedik ki az embernél és majmoknál is jólismert nyelvcsap (uvula). Azon a helyen, ahol az ínyvitorla leszáll a nyelv gyökeréhez, találjuk azokat az ugyancsak jól ismert nyirokcsomó-halmazokat, melyeket mandolánaknak (tonsillae) nevezünk; a mandolák tulajdonképpeni jelentőségét nem ismerjük, valószínű feladatuk a betegségokozó anyagok megkötése.
A gyomor (ventriculus) és bél (intestinum) az emlősöknél nagyobb ürtartalmú, a test hosszához viszonyítva a belek is hosszabbak és terjedelmesebbek, mint a madaraknál. Az emésztés voltaképpeni szervének ez a szerkezete nyomban elárulja, hogy az emlős állat egyszerre nagyobb mennyiségben veszi fel táplálékát, egyszerre tölti meg gyomrát, (mely a növényevőknél a táplálék tápanyagszegénysége következtében több részre tagozódás útján szerfölött megnagyobbodhatik, mint a kérődzőknél) azután pihen és emészt és végül ürülékét is ritkább időközökben s egyszerre nagyobb mennyiségben rakja le; a madár ellenben úgyszólván folyton eszik, folyton mozog és folyton ürít.
A gyomor, melybe a táplálék a nyelőcsövön (oesophagus) át kerül, az emlősöknél – kevés kivétellel (például a fóka) – harántul fekszik és balra jobban kiöblösödik (curvatura major), mint jobbra (curvatura minor). Ezenkívül különféle kiöblösödések is felléphetnek, sőt a kérődzőknél, mint ismeretes, tökéletesen négy részre tagolt gyomrot találunk. A gyomor sokféle alakja mindig kettős feladatára – emésztés és táplálék felhalmozás – vezethető vissza. Még a külsőleg nem tagolt emlős-gyomor fala is sok esetben belülről tagolt, olyképpen, hogy van mirigyes és mirigytelen része is, jeléül annak, hogy a munkamegosztás elve ilymódon érvényesült. A gyomornál a gyűrűs izmokkal ellátott gyomorcsukló (pylorus) vezet a sokszorosan kígyódzó vékonybélbe (intestinum tenue), amely a felszívó bolyhok gazdagságával tűnik ki. A vékonybélben szívódnak fel a folyóssá vált ételpépből a tulajdonképpeni tápanyagok és pedig a bélbolyhok segítségével.
A megevett tápláléknak csak azon részeit használhatja fel a szervezet testének felépítésére, amelyet vízben oldható anyagokká képes feldolgozni. A szénhidrátoknak – lisztes ételek keményítő tartalmának (kenyér, tészta stb.) – megfelelő szétbontása már a szájban megtörténik, a fehérjék (hús) feldolgozását pedig nagyjában már a gyomornedvek elintézték. Mindazonáltal maradnak még feldolgozatlan szénhidrátok és fehérjék is, sőt teljesen változatlanul jutnak be a bélcsatornába a különféle zsírok. Ezen még változatlan anyagok feldolgozását a bélcsatornába szájadzó különféle járulékos mirigyek váladékai végzik.
Így a hasnyálmirigy (pancreas) váladéka többféle hatóanyagot tartalmaz, amelyek közül a trypsin a fehérjére, a diastaze a keményítőre és a steapsin a zsírokra hat átalakítóan, s így váladékával az emésztést nagyban elősegíti.
Az emlős test legnagyobb igazi mirigyei a máj (az elefánt májának súlya csaknem egy métermázsa), mely a hasüreg jobboldalán közvetlenül a mellkas alatt van. Összetett, csöves mirigy, mely az epe kiválasztásán kívül nagy szerepet játszik tartalék tápanyagok felhalmozásában is; ezenkívül eltávolítja a szervezetből a káros és mérges anyagokat. Éppen ezért nem tekinthető egyszerű emésztési mirigynek. Váladéka a sötétzöldesbarna epe keserű ízű, kellemetlen szagú folyadék; hatóanyagai a bélbe jutott táplálék zsírját elszappanosítják, vízben oldhatókká és felszívódásra alkalmassá teszik. Az epe a bél mozgására is élénkítően hat és a béltartalom rothadását is gátolja.
Említettük, hogy a máj nagy szerepet játszik a tartalék tápanyag felhalmozásában is és mint ellenőrző szerv is szerepel. Mindkét folyamat olyképpen történik, hogy a vér a verőceéren (vena portae) át a gyomorból és a belekből a májba jut, – még mielőtt a szívbe jutna, tehát a máj ellenőrző szervként van beiktatva a vénás áramba, sőt a középponti erek az elhasznált vért is a májba vezetik – és a máj sejtjei egyrészt a bélből odaszállított szénhidrátokból májcukrot, glikogént készítenek, másrészt a vérben levő mérges anyagokat lekötik; az így ellenőrzött vér végeredményben a hátulsó üres vénába (vena cava caudalis) s innen a szívbe jut. A glikogén beraktározásának ez a módja rendkívül emlékeztet a növények keményítő felhalmozására; a glikogén egyébként olyan vegyület, amely rokon a keményítővel és a keményítőcsirizzel vagy dextrinnel. A májban a májcukron kívül elég nagy mennyiségű zsír is halmozódik fel és pedig apró golyócskák alakjában. Hízlalással a zsírfelhalmozás a májban is fokozható.
Ahol a vékonybél a vastagbélbe (intestium crassum) nyílik, ékelődik be a bélcsőbe a vakbél (caecum). A vastagbélbe jutott anyagból a folyadék egy része felszívódik, a többi bélsárrá sűrűsödik be; az emészthetetlen részek megfelelő rögökké idomulnak és kiüríttetnek. A vakbél a növényi rostok, cellulose, emésztésében játszik nagy szerepet és éppen ezért a növényevőknél igen nagy, a húsevőknél pedig teljesen hiányzik.
Természetes, hogy egy ilyen hosszú és mégis aránylag kis helyen – a hasüregben – elhelyezett szervnek, mint aminő a bélcsatorna – mely például a szarvasmarhánál a test hosszának mintegy húszszorosa (58 m) – különösen célszerű elrendezésre és megerősítésre van szüksége, hogy ne tolódjék el túlságosan a helyéről. A belek megerősítésére a hasüreget kibélelő hashártya kettőzetei (duplicaturae) szolgálnak, amilyenek a savós szalagok, a bélfodor (mesenterium) és a cseplesz (omentum). A bélcsövet a fodros alakjáról elnevezett bélfodor tartja felfüggesztve, míg a zsírsávokkal átszőtt hálószerű cseplesz kötényszerűen lóg a has belei előtt.
Már a vérkeringéssel és a rekeszizommal kapcsolatban említettük, milyen nagy különbség van az emlősök és madarak légzőszervei között. Ugyanilyen különböző – mind a két gerinces osztályban – a hang képződésének módja és helye is. Minthogy az emlősállatnak megközelítően nincs olyan hangja, mint a legtöbb madárnak, azért hangszervei is sokkal egyszerűbb alkotásúak. Ahol a légcső kétfelé ágazik és a hörgők (bronchusok) a tüdő felé indulnak, van a madarak alsó gégefője, ahonnan a csattogó madárdal fakad; az emlősöknél nyoma sincs egy második gégefőnek. Az emlősök hangja a légcső felső kezdetében, az ismert gégefőben (larynx) keletkezik, ahol a különféle módon elhangzott és kifeszített hangszalagok (ligamenta vocalia) rezgése útján jön létre. A gége üregében, körülbelül ennek a közepetáján, a nyálkahártya az oldalfalakról két redő alakjában beemelkedik, ezek közül a gégebejáratához közelebb eső a tasakredő, a mögötte levő a hangredő, amelyben a hangszalag van; a két redő között találjuk a hasíték alakú hangrést (rima glottidis). A gége porcai között különféle tasakszerű bemélyedések is lehetnek, melyek a rezonancia szervei s így a hang erősítésére szolgálnak (bőgő majom). Az emlősök gégefőjén még egy jellemző dolgot találunk, nevezetesen a gégefedőt (epiglottis), mely nyelés közben előlről ráfekszik a hangrésre, míg a többi gerincesnél izomerővel záródik. Újabban, minthogy ízlelési bimbó van rajta, ízlelőszervnek is tartják a gégefedőt.
A gégecső (trachea) a gége közvetlen folytatásaként húzódik a nyakon a mellkasba; hossza a nyak hosszától függ; szabadabb mozgása miatt vázát porcos gyűrűk alkotják, melyek nem mindig alkotnak teljes gyűrűket, a légcső nyitvatartására szolgálnak, és összenyomását is meggátolják. Alsó végén, a tüdőszárnyaknak megfelelően két ágra, a hörgőkre oszlik, melyek a tüdőn belül tüdőfává ágaznak el. A tüdőfa legvékonyabb ágacskái, számos üres duzzadással, az úgynevezett tüdőhólyagocskával (alveola) vannak megrakva, melyeknek fölötte vékonyfala sűrű hajszálerekkel van átszőve; a tulajdonképpeni lélekzés, azaz a gázcsere a vér és levegő között, az alveolákban történik.
Az emlősállat tüdeje abban különbözik a madárétól, hogy egészen szabadon fekszik a mellkasban. Kivételesen (elefánt) a tüdő összenőhet a mellkast kibélelő mellhártyával (pleura); ilyen nem kívánatos összenövés valamilyen betegség következménye is lehet. A tüdő a madaraknál a mellhártyával nőtt össze s a tüdővel összefüggő légzacskók a repülő madaraknál nagy szerepet játszanak a testsúly csökkentésében. Az emlősöknek nincsenek légzacskói, még a repülő denevéreknek sem.
Az emlős tüdő finom és legfinomabb szerkezetét – igen gazdag és változatos anyagon – rendkívüli alapossággal tanulmányozta Schulze berlini zoológus. Vizsgálatai, melyek e téren bizonyos tekintetben befejezték e szerv kutatását, itt teljes egészében nem közölhetők. Ám egyik fölötte tanulságos eredményét meg kell említenünk. E szerint a tüdő belső szerkezete és az élethez szükséges energiák – életerő – között szoros összefüggés van, még az emlősök osztályán belül is. Így Schulze szerint a macskatüdő körülbelül 400,000.000 alveolát tartalmaz. A vele körülbelül egyenlő nagyságú háromujjú lajhárnak azonban csak 6,250.000 alveolája van. Az alveolák különböző nagyságát is figyelembe véve, kiszámíthatjuk a légzésben részt vevő – respiraciós – felületet; az eredmény a macskára vonatkozólag 20 m2, a lajhárra vonatkozólag csak 5 m2. Schulze „hajlandó ezt a különbséget arra vonatkoztatni, hogy a macska, mit apró ragadozó, nagyon erős izomtevékenységet fejt ki, ezért felette élénk az anyagcseréje, s így nagy lélegző felületre van szüksége; míg sarlóalakú karmaival a fa ágain lógó és minden megerőltetés nélkül leveleket fogyasztó lomha lajhár, csak kevés mozgás végez s ezért anyagcseréje sokkal kevésbbé élénk, mint a macskáé; így tehát jóval csekélyebb lélegző felületre van szüksége, mint amannak”.
Hausmann régebbi megállapítását, hogy szomszédos tüdőhólyagocskák válaszfalai áttörnek, Schulze végérvényesen igazolta és érdekes módon hozta ezt is összefüggésbe a különböző emlős csoportok különböző életenergiájával. Erről a következőket mondja: „A tüdőhólyagocskák válaszfalában a legtöbb áttörést egyes rovarevőknél találtam, így különösen a sünnél, a vakondoknál és a cickányoknál, főképpen azonban a legkisebb német cickánynál (Sorex minutus). Magától értetődik, hogy az alveola válaszfalainak is nagymérvű átlyukgatása – perforációja –, nagy jelentőségű a gázcsere fokozásánál, amennyiben itt, a csak vékony hártyával körülvett hajszáledényeket, csaknem minden oldalról éri a levegő, míg más emlősöknek csak kevéssé átlyukgatott válaszfalaiban, a hajszálerek csak két egymással szemben fekvő oldalon érintkeznek a levegővel. Hogy ez a fokozott gázcsere-alkalom épp azoknál az állatoknál található, melyeknek különösen intenzív az anyagcseréje, az érthetőnek látszik. Hiszen a vakondoknak naponta körülbelül annyi állati táplálékra van szüksége, mint amennyi saját testének súlya. Hasonlóképpen a cickányok is, mint mindenki tudja, a legfalánkabb emlősök közé tartoznak. Hacsak legkevésbbé éheztetjük őket, elpusztulnak”.
A vese (ren) feladata, hogy a testből az anyagcsere folyékony bomlás termékeit kiválassza; már külső alakjában is nagyon eltér a madarakétól. Az emlősvesének rendesen egész határozott és hajlott körvonalaival nagyon jellemző alakja van; a madárvese alakjáról aligha beszélhetünk, minthogy az a keresztcsont mélyedéseibe van bepréselve. A kiválasztott vizelet külön vezetők (uretheres) útján jut a vesékből a húgyhólyagba (vesica urinaria), ahonnan időnként eltávolodik.
A szaporodás szervei két külön ivarra oszlottak el. A női ivarszervek leglényegesebb sajátságait – melyek származástani és rendszertani szempontokból fontosak – már a fejlődés kapcsán ismertettük. Meg kell még említenünk azt is, hogy rendesen mind a két petefészek egyformán van kifejlődve, azonban a kloákások egyik petefészke csökevényes, éppúgy, mint a madaraknál. A hím ivarszerveknél az a legfeltűnőbb, hogy a legtöbb emlősnél a hímcsíramirigyek nem maradtak a hasüregben, hanem állandóan, vagy legalább az ivarzás idején a hasüregen kívül levő bőrkitüremkedésben foglalnak helyet. Aki részletesebb felvilágosítást óhajt, ebbeli kívánságát bármelyik anatómiai kézikönyv vagy egyetemi tankönyv útján kielégítheti.
Mielőtt az emlősök állati életének leírásával foglalkoznánk, szólanunk kell még egyetmást az idegrendszerről és az érzékszervekről. Hiszen ezek azok, amelyek a fent leírt testnek „életet adnak” és vonatkozásba hozzák a külvilággal. Újabban több különböző érzéket tételezünk fel, így például külön hőmérséklet, nyomás és fájdalomérzéket, mert pontos kísérletek azt bizonyítják, hogy a bőrnek csak bizonyos helyei, az úgynevezett nyomás- és fájdalompontok közvetítik a fentnevezett érzéseket.
A közismert öt érzék közül, egy, a tapintási érzék, az emlősöknél az egész test felületén található, és feladata, hogy a külvilág tárgyaival való érintkezésről értesítsen. A bőrben mindenütt vannak tapintó sejtek (Merkel-féle sejtek), itt-ott tapintó testecskék is (Pacini-féle Meissner-féle tapintótestecskék és végtestecskék), melyeknek közös tulajdonsága, hogy vagy rajtuk, vagy bennük egy idegágacska végződik, mely többé-kevésbbé hurokhálókat képezve, a legfinomabb rostokra oszlik. A testnek azon a helyein, amelyek gyakran jönnek érintkezésbe idegen tárgyakkal – mint ajkak és orr, a kéz és láb ujjainak hegye, tenyér és talp – tapintósejtek és tapintótestecsék halmozódnak fel. Az ajkakon többnyire a megérzés szolgálatában álló tapintószőröket is megtaláljuk, melyeknek szőrtüszője többnyire vérrel telt üregeket tartalmaz és idegvégződésekben is gazdag (sinus-szőrök). Valószínű, hogy ezek a tapintószőrök – amelyek elsősorban a ragadozóknál, rovarevőknél és rágcsálóknál fordulnak elő – arra valók, hogy az állatot figyelmeztessék arra, vajjon a sűrűségeken és szűk nyílásokon átférnek-e?
Míg a tapintás érzékének általános elterjedését tekintve, nagy a hasonlósága a madarak és emlősök között, addig ízlés dolgában – a tápláléknak a nyelvvel való megvizsgálását értve alatta – a két gerinces osztály igen különböző. Már az elszarusodott madárnyelvből arra következtethetünk, hogy a madár táplálékát inkább tapintással vizsgálja meg. Más megfigyelések is azt bizonyítják, hogy a madár életében íz és szag sokkal kisebb szerepet játszik, mint az emlősöknél. Az emlősöknél az ízlelés székhelye a nyelven és a szájpadláson van. Ez az érzék tehát szűkebb területen fordul elő, de még mindig nem különálló szerv, mely kizárólag az ízlelés szolgálatában állana; érzeteit az ízlési kelyhek és bimbók közvetítik, melyek úgy a lágy szájpadláson, mint a nyelvtövén vannak szétszórva; legsűrűbben a körülárkolt és leveles szemölcsök oldalfalában találhatók.
Mint ahogy az ízlés csak a folyékony és nyálban oldott tápanyagok tulajdonságairól vesz tudomást, úgy a szaglás sejtjeire csak gázalakú anyagok hatnak. A szaglási szervnek, az orrnak, a levegő állandó ellenőrzése a legfontosabb feladata. Az emlősök szaglóérzékének azonban más feladatai is vannak. A szaglás az emlősállatoknál sokkal nagyobb jelentőségű, mint a gerincesek bármely osztályánál, amit a szervnek nagy kiterjedése is elárul. Nagy részüknél a szaglás az alapérzék, ez irányítja minden cselekedetüket, erre építik egész érzéki és lelki életüket. Az orr üregét az orrsövény két félre osztja, mind a kettőbe az oldalfalakról két orrkagyló (néha három) emelkedik be, melyek három orrjáratot különítenek el. Az orrüregnek a rostacsonttal (os ethmoidale) szomszédos részében az orrsövényt és a felső orrkagylót, a szagló-nyálkahártya borítja, melyben szaglósejteket találunk. A szaglóideg (nervus olfactorius) rendesen nem egy idegből áll, hanem az agyvelő szaglólebenyéből, a rostacsont számtalan lyukacsain áthaladó és végeredményben ecsetszerűen elágazó szaglórostok összességét értjük alatta; megemlítésre érdemes, hogy a madarak és a csőrösemlős rostacsontján a szaglóideg részére csak egy-egy nyílás van. Az alsó orrjárat a lélekzőjárat a legtágasabb, a hortyogókon keresztül a garatba nyílik. A középső szűk orrjárat erősen csavarodott orrkagylóival, leginkább szűrőhöz hasonlítható, mely az orrba futó levegőt egyrészt megtisztítja a portól és szennytől, másrészt felmelegíti azt, ami a hirtelen lehülés és meghülés ellen védi a szervezetet. Ezért igen erősen fejlett orrkagylókat azoknál az állatoknál találunk, amelyek hidegben élnek (fókák), vagy azoknál, amelyek orrukkal a földet érintik; így a rövidlábú rágcsálóknál és a folyton szimatoló ragadozóknál. Az emlősök között egyedül a ceteknek nincs szagló idegük. A cetek orrjárata tehát csak légjárat, amelynek külső nyílásai a tengeri utazóktól oly sokat emlegetett fecskendőlyukká olvadtak össze.
Az orr néha orrmánnyá nyúlik meg (például disznó, elefánt) s ilyenkor a táplálékszerzés (túrás, megragadás) szolgálatába szegődik, ami korántsem oly csodálatos, mint első pillantásra látszik, hiszen a táplálékszerzésben a szaglásnak igen nagy szerepe van.
A hallás készülékével kapcsolatban meg kell emlékeznünk arról, hogy az emlősöknek – ellentétben a madarakkal – három hallócsontocskájuk van, amelyekről a koponyával kapcsolatban már megemlékeztünk.
Ezenkívül az emlősökre rendkívül jellemző még a belső fülben található csiga (cochlea) alakja is – a csigában van a hallóideg –, mely az emlősöknél csavarodott, míg a madaraknál (lagena) tompa kúpalakú. A madaraknál hiányzik a külső fül is, míg az emlősöknél jól fejlett s izmok által mozgatható fülük van, melynek alakja – főképpen ha tölcséralakú, mint a patásoknál – már messziről elárulja azt, hogy az a hang felfogására szolgál. Henneberg (Giessen) legújabban a fülkagyló alakjából olyan berendezésekre következtet, amelyek az emlősök őseinél, sőt részben a ma élőknél is, egyrészt a víznek, másrészt a különféle szennynek a fülbe való hatolását gátolják meg. A kiálló fül azonban nem bizonyítéka annak, hogy a hallás az emlősök életében nagyobb szerepet játszik, mint a madarakéban, amelyeknek bár nincs fülük, igen kitűnően hallanak. Az emlősök közül külső füle csak néhány földalatti és viziéletet élőnek nincs; ezek életelemükben, a földben és a vízben, a hanghullámokat egész testükkel fogják fel.
A látás az emlősállatnál közelről sem olyan feltétlenül uralkodó alapérzék, mint a madaraknál, ahol van rá eset, hogy a szem olyan nehéz és olyan nagy, mint az agyvelő, a legvégsőkig keresztül vitt takarékossági elv ellenére, amely a madár testének felépítésében annyira érvényesül. Vannak földben élő – ásó – emlősök, amelyeknek szeme egészen elcsökevényesedett, de egyetlenegy emlősnél sem találunk olyan berendezéseket – amelyek a szemnek különböző távolságokra való gyors beállítást tennék lehetővé, amelyek a szemet oly bőségesen ellátnák táplálékkal –, mint a madaraknál, melyeknek szemét e tulajdonságot általában jellemzik. Mindazonáltal, a legújabb kutatások szerint, az emlősök is élesen látnak, meg tudják látni a legkisebb és legfinomabb mozdulatokat is és pedig sohasem sejtett mértékben, mely messze felülmúlja a vadászok erre vonatkozó tapasztalatait. A macska szemének villogása, melyhez hasonlót más, homályban jól látó emlősöknél is tapasztalhatunk, úgy keletkezik, hogy a szem érhártyája (chorioidea) – a látóideg belépése fölött félkör vagy háromszög alakjában finomabb hullámos rostokkal – fénylő kárpittal van kibélelve, mely a beeső sugarakat visszaveri, s így azokat az előtte levő fényérzékeny recehártyán (retina) másodszor is átküldi.
A szemek egymáshoz való helyzete a különböző emlőscsoportokban más és más. A madaraknál és a többi gerincesnél rendesen az egyik szem nem látja azt, amit a másik szem lát, vagyis látómezőik tökéletesen elkülönítettek. Az emlősöknél az egészen vagy részben elkülönített látómezőktől, az egységes látásig – így lát az ember és a majom – minden átmenetet megtalálunk. A pislogó hártya, mely a madárnál mint harmadik szemhéj lép fel és a belső szemzugból kiindulva fedi a szemgolyót, az emlősöknél – mint nem mozgatható félholdalakú redő (plica semilunaris) – csökevény alakjában van meg. A szem segéd- és védőszervei közül, az emlősökre jellemző, még a szemhéjak szőrzete, a szempillák is.
Az egész állati élet központi szerve: az agy- és gerincvelő a főidegtörzsekkel, világosan mutatja, hogy az embernek – ennek a gondolkodó és beszélő-emlősnek – az emlősállatok a legközelebbi rokonai, az „állat” szót olyan értelemben véve, amilyenben már Aristoteles használta, mikor az embert „Zoon politikon”-nak, azaz társas, államképző állatnak nevezte.
Minden magasabb és finomabb életműködés kimutatható székhelye a nagyagy; fejlődése a halak igénytelen előagyából indul ki, amely a két agy féltekének (hemisphaeria cerebri) mind hatalmasabb kifejlődése által – már a többi gerinces osztályokban is lassanként előtérbe nyomuló – nagyaggyá lesz; fejlődésének legmagasabb fokát, az emlősök osztályán belül éri majd el, ahol a két agyfélteke nemcsak legnagyobb része az agyvelőnek, hanem legelőbbre is helyeződött, olyképpen, hogy az összes többi agyrészletét is befedi. Legfontosabb részük az úgynevezett szürke agykéreg felülete is bámulatosan megsokszorosodott, úgyhogy ráncokba gyűrődött; így keletkeznek a barázdákkal (sulci) elválasztott agytekervények (gyri), amelyeknek fejlettsége azonban egymagában még nem a tökéletesedés jele, mert például a majmok között vannak síma agyfelületűek, míg a patások és cetek agyveleje csaknem olyan tekervényes, mint az emberé; a szellemi működésre szolgáló kéregrészlet, a homloklebeny, azonban ezeknél jóval kisebb, mint az embernél.
Minden életműködés központja – székhelye és kiindulópontja – az agy egy egészen határozott helyén van minden életműködésben zavar lép fel akkor, ha betegség támadja meg, vagy kísérlet céljából műtéttel távolítjuk el az életműködésnek megfelelő agyrészletet. Egy élettani tankönyvből vett néhány példa jobban megvilágítja a mondottakat. A mótorikus, mozgatóközpontból az inger vezetés útján jut el a megfelelő ideghez, amely ingerlékenységénél fogva az ingerre reagál és a szükséges mozgást kiváltja. A tankönyv szerint külön központ irányítja a kutya nyakszirtizmainak, a mellső végtag hajlító és mozgató, a hátsó végtagnak, arcizmoknak, a farok csóváló, az állat haladó, szemhéjak csukódó, a nyelvnek, az állkapcsok záródó stb. mozgását. A tudatos érzéki észrevételeknek ugyanilyen jól kifejlett és helyhez kötött érző – sensoricus – központjai vannak az agyban ahová az érző és érzékelő idegek vezetik az ingert. Így érthető, hogy például az ember, vagy a kutya stb. vak lehet vagy megvakítható, jóllehet, szeme minden hozzátartozó szervvel együtt teljesen ép és egészséges, még az agyba vezető látóideget is beleértve.
Csak egyetlen olyan központot ismerünk, amelyet csak az embernél találunk meg, amely minden más állatnál – legyen az bár emlős – hiányzik; ez az úgynevezett Broca-féle központ – nevét felfedezőjétől, a tudományos embertan atyjától kapta –, mely a tagolt beszédnek agyközpontja. Ez a helyes magyarázata annak, hogy tulajdonképpen hol és mi módon keletkezett a beszéd, s miért nem szabad csodálkoznunk azon, hogy míg az ember beszél, jóllehet a beszédnél szereplő szervek (gége, nyelv, ajkak) aránylag egyszerűek, addig az állatok, még az emberszabású majmok sem beszélnek, noha ugyanezek a szervek náluk sokszor finomabb felépítésűek és többfélék is, és mellékszervekkel is bőségesebben felszereltek. Legalább is tulajdonképpeni, magasabb értelemben vett beszédjük nincs, különféle kedélyhullámzásaikat csak különféle hangokkal tudják kifejezni.
Természetes, hogy az egyes agyközpontok nagysága és a benne összefutó idegrostok száma az állat növekvő testnagyságával együtt nő, a koponya nagysága és növekedése azonban egészen más körülménytől függ. Éppen ezért az általában nagytestű emlősöknél – amelyeknél a koponya növekedése valamilyen oknál fogva korlátok közé szorított, mint már említettük – az agy összegyűrődni kénytelen és így tekervényeket alkot, a nélkül, hogy ez az állat intelligenciáját növelné, vagy azzal bármiféle összefüggésbe hozható volna; az agytekervények száma tehát önmagában nem kritériuma a szellemi magasabb rendűségnek.
Kétségtelen és ismeret, de még egyáltalában nincs megmagyarázva az emlős idegeinek az a sajátsága, hogy az idegek kereszteződnek egymással, másrészt minden agyfél az ellenkező testféllel van összefüggésben; így például baloldali agyvérzés a jobboldal bénulását vonja maga után.
Könnyebben érthető, hogy az emlősök agyveleje csecsemőkorban aránytalanul nagyobb, mint kifejlett korukban. Weber az agyvelő súlyát – nagyon helyesen – a testsúlyhoz viszonyítja; igen érdekes adatokat közöl – már említett könyvében – az oroszlán agyvelejének és testének súlyviszonyairól; így öthetes korban a kettő úgy viszonylik egymáshoz, mint 1:18, négyhónapos korban 1:80, tizenegyhónapos korában 1:184 és végül öreg korában 1:546. Rendkívül érdekesek Webernek azok az adatai is, amelyek az egy nembe tartozó, különféle nagyságú fajok – de kifejlődött állatok – agyvelejének súlyáról tájékoztatnak; így a törpe macska arányszáma 1:56, a leopárdé 1:168, az oroszláné 1:546. A példának említett számok azt bizonyítják, hogy az emlősök érzéki és lelki életéhez, egy bizonyos minimális agyvelő szükséges; ha ezt a minimális agyvelőtömeget a faj megszerezte, s így a legszükségesebb agyvelőközpontok kifejlődhettek, az agynak tovább gyarapodnia nem okvetlenül szükséges.
Az élő és kihalt emlősök agytömegét is össze lehet hasonlítani egymással, akkor, ha a kihalt állatok agytokját gipsszel kiöntjük; a gipszöntvény ugyanis az állat agyvelejének pontos másolata. Minden esetben bebizonyosodott, hogy a kihalt állatok agyveleje jóval kisebb, mint a ma élő emlősöké. Ez már önmagában is bizonyítja azt a nagyon valószínű feltevést, hogy az emlősök a törzsfejlődés folyamán nemcsak testi, hanem szellemi fejlődésen is keresztül mentek; a szellemi fejlődés legtökéletesebb fokát az ember képviseli. Kétségtelen, hogy az emberi és az állati szellem között óriási különbségek vannak; kétségtelen továbbá, hogy a ma élő emlősök és az ember között testileg sincs meg az összekötő-kapocs – a missing link –, a hiányzó láncszem. Azonban egyik éppoly kevésbbé csodálatos, mint a másik, hiszen fel kell tételeznünk, hogy ez a lény – sem ember, sem állat – nem maradhatott meg a földön fejlődés nélkül. Hiszen el kell ismernünk, hogy a ma élő állatok – sőt még az ember is – nem egyebek, mint sokszor igen bámulatosan célszerű „alkalmazkodások” bizonyos életkörülményekhez, amelyek fölötte ügyesen, hogy ne mondjam, agyafurtan használják ki az élet bizonyos lehetőségeit. Ha ezekkel szemben a kihalt „özönvízelőtti” állatokat nézzük, a ma élők mellett legtöbbjük az „esetlen szörny” benyomását kelti. A kihalt állatok környezetének ismerete még inkább fokozza azt az érzésünket, hogy a szélsőséges – kicsapongó fantáziából származott – formák nem voltak célszerűek s így magukban hordták pusztulásuknak csíráit. A kevésbbé szélsőséges, igénytelenebb, s éppen ezért harmónikusabb formák pedig tovább fejlődtek és ezekből kell levezetnünk a ma élő emlősöket. A fejlődés természetesen nem egy irányban haladt előre, hanem lehetőségekhez mérten. A fejlődés különböző irányait a csoportosítás lehetőségeiből is kiolvashatjuk, természetesen csak az esetben, ha különböző szempontok szerint csoportosítjuk az emlősöket. Így mozgásuk alapján megkülönböztetünk: futókat, ugrókat, úszókat, kúszókat és röpködőket; a mellső láb szerkezete alapján: karmosak, patások, úszólábúak és halhoz hasonló (cetek) emlősállatok; táplálékuk szerint lehetnek: hús-, hal-, dög-, növény- és minden-evők; vannak ragadozó, rágcsáló, foghíjas, kérődző, orrmányos és stb. emlősök.
Minthogy az ember testileg az emlősállatok közé tartozik, némelyek ezeket a szellemi tevékenységek szempontjából is az ember közelébe és melléje állították, jóllehet ez a felfogás az elfogulatlan megfigyelés és a tudományos kritika előtt nem állhat meg. Ezzel a felfogással gyökeresen végezni kell.
Wundt a lipcsei filozófus „Vorlesungen über die Menschen- und Tierseele” (Előadások az emberi és állati lélekről) című művében a következőket mondja: „Ha pontosabban megvizsgáljuk mindazt, amit hiteles megfigyelések igazolnak, ha továbbá azt a takarékossági elvet követjük, mely szerint bonyolult, magyarázó okokat csak akkor szabad igénybe vennünk, mikor az egyszerűek be nem válnak, akkor az állatok egész szellemi életét visszavezethetjük egyszerű képzettársulási (asszociáció) törvényekre, míg azok a bélyegek, melyek a valóságos eszmélkedésre és az értelemnek vagy képzelőtehetségnek cselekvő tevékenységre jellemzők, teljesen hiányzanak”. Heck ugyanezt más helyen, más szavakkal így mondja: „Eddig még egyetlen esetet sem állapítottak meg, mely azt bizonyítaná, hogy valamely állat szellemi tevékenysége a fölé a színvonal fölé emelkedett volna, melyet a pszichológus képzettársítás néven ismer; ez az érzéki észrevételek és tapasztalatok célszerű összekapcsolása cselekedetekkel, többnyire úgy, hogy a cselekvés az állat egyéni jóllétének vagy faja fennmaradásának javára szolgál”. Ezek az asszociációk azonban teljesen külsőségesek, nem alapszanak az ok és okozat belső összefüggésének meggondolásán vagy igazi belátásán. Ezt északamerikai kutatók, kortársaink, hosszú, tervszerű kísérletsorozatokkal igazolták. Mindenféle kísérleti állat (patkány, macska, majom) csak úgy jutott táplálékhoz vagy szabadsághoz, ha egy bizonyos, többé-kevésbbé hosszadalmas utat megtalált, vagy egy bizonyos gépezetet mozgásba hozott. Ezt hosszabb-rövidebb idő alatt valamennyien megtanulták; de mindig fel lehetett ismerni, hogy eleinte teljesen terv nélkül próbálgattak, és e közben véletlenül találták meg a helyes utat. Ezáltal meg volt adva a helyes asszociáció, amely annyira megszilárdult, hogy csekély gyakorlat után, a kísérleti állatok újabb kísérleteknél, azonnal célszerűen cselekedtek. Sohasem történt azonban, hogy egy kísérleti állat cselekvés előtt értelmesen megfontolta volna a dolgot és miután a helyes megoldást megtalálta, első cselekvésével a célszerű megoldást találta volna el. Tehát még ahol ember és állat egyformán, célszerűen cselekszenek is, különböző utakon érnek oda: az állat, még a fejlődés magasabb és legmagasabb fokán álló emlősállat is, véletlen, az emlékezetben tartott tapasztalat útján, az ember pedig értelmes megfontolás és az okozati összefüggés valóságos belátása útján. Vannak esetek, amikor állati és emberi cselekvések annyira egyformák, hogy más megoldás szinte elképzelhetetlen, mint az, hogy az állatnak emberi szellemiképességei volnának; mégis, igazán tudományos kutatások ilyenkor is arra az eredményre vezetnek, hogy ezekben az esetekben is egészen más összefüggésekről és keletkezési módokról van szó. Klasszikus példája ennek a „bölcs Hans”, a berlini Osten úr „tudós” lova, amelyet gazdája mindarra megtanított, amit egy népiskolai tanulónak tudnia kell: írásra, olvasásra és számolásra; a tanítás módszere állítólag ugyanaz volt, amit a népiskolákban a gyermekek tanításánál alkalmaznak, csak az állat némaságának megfelelően módosítva. Erre vonatkozóan Heck a következőket mondja: „A bölcs Hans esete megtanított arra, hogy az állatok oly szellemi ténykedésekre képesek, melyek látszólag hajszálnyira hasonlítanak a sajátlagosan emberi cselekvésekhez, valójában azonban egész más módon keletkeztek. A „bölcs Hans” látszólag éppúgy számolt és olvasott, mint egy népiskolai tanuló, pedig valójában semmit sem tudott számokról és betűkről, hanem csak feszülten figyelt az előtte álló ember legkisebb, öntudatlan mozdulataira, amelyek jelezték, hogy mikor kell patáinak abbahagynia a kaparást (ilyen, igen ügyes rendszer helyettesítette a „tanítás”-nál a beszédet), hogy répát és kenyeret kapjon”. Pfungst Oszkárnak, a berlini pszichológusnak érdemes, hogy ezt felfedezte és kísérletekkel bizonyította. „Gondoljuk csak el”, folytatja Heck, „hogy ezután a tapasztalat után mily kételkedéssel kell fogadnunk az állatok rendkívüli intelligenciájának látszólag megcáfolhatatlan bizonyítékait, melyekről az irodalom beszámol!” Sokszor nyilvánvaló, hogy valamely cselekvést fölösleges módon túlbecsülnek. Mindenki ismeri az elefántnőstény történetét, mely borjúját a veremből ki akarja menteni. „Hűségesen kitart mellette, mígcsak a vadász közeledése el nem űzi. A verem fenekén sok földet és gallyat találnak és ebből minden további nélkül arra következtetnek, hogy az anyaállat mindezeket azért dobta a verembe, hogy borjújának a kimászást lehetővé tegye, holott sokkal közelebb fekszik az egyszerű, csaknem magától értetődő feltevés, hogy a föld és az ágak egyszerűen beleestek a verembe, amikor az anyaállat a gödör szélét taposta, folytonosan próbálgatva, hogy borjúját orrmányával kiszabadítsa”. Sok olyan eset is van, mikor szinte reménytelennek látszik, hogy más magyarázatot találjunk, mint a közkedvelt feltevést, mely szerint a magasabbrendű emlősök úgyszólván emberek, beszélőképesség nélkül. Ily esetekben a dolog mélyére kellene hatolnunk: gyakran elbámulnánk az eredményen! Így például egy előkelő német családi folyóirathoz beküldtek egy hosszú, csodálatos, megható történetet, mely egy farkas és tengeri malac közti barátságról szólt (a cikkíró nő volt!). Heck megvizsgálta az esetet s kutatásának eredménye az volt, hogy a történetből egyetlen szó sem volt igaz.
Valószínűen hasonló elbírálás alá esik az a sok állatbarát, akinek túlzott, mindazonáltal szilárd meggyőződése, hogy kedvencének „emberi értelme” van. Sok állatbarát öntudatlanul kiszínezi, túlozza, amit kedvence tesz. Minden józan megfigyelő, minden önállóan ítélő kutató, ez állatbarátok szemében „szeretetlen és meg nem értő”. Mégis legfőbb ideje, hogy elfogulatlanabb nézetekre tegyünk szert az állat-, de főkép emlőslélektanban, nemcsak a tudósok, hanem az állatbarátok körében is.
Ha meg akarjuk ismerni az állatok szellemi tevékenységének valódi természetét, úgy a logikán és a pontos tudományos kutatáson kívül az az anatómiai és főkép agy-anatómiai vizsgálatokat sem mellőzhetjük. Erre vonatkozik Edinger (Frankfurt a. M.) előadása, melyet „Az összehasonlító anatómia viszonya az összehasonlító psychologiához” címmel a kísérleti psychologusok III. kongresszusán tartott. Előadásának végén a következőket mondja: „Ami azonban minden állatot az embertől megkülönböztet, az a neencephalon nagysága (Edinger neencephalonnak azokat az agyrészeket nevezi, melyek a magasabb szellemi tevékenységeknek szolgálnak, elsősorban a nagy agyvelőt két hemisphaeráját). Egy óriási gorillának kisebb agyveleje van, mint az ember-csecsemőnek. Az ember valósággal megdöbben, ha kiveszi a koponyából a kicsinyke agyvelőt, amelyen – eltekintve a hátsó és középső részlet csekélyebb fejlettségétől – a homloklebenyek is hiányzanak. Ezek a homloklebenyek különböztetik meg elsősorban az embert az állattól. Az emberi kórtan (ebben az esetben az agybetegségek tudománya, tekintettel a vele összefüggő szellemi zavarokra) arra enged következtetni, hogy a homloklebenyek éppen a magasabb lelki működéseket, az elvonásokat (absztrakció) a fogalomképzést teszik lehetővé. Nyilván csak a beszélőképességgel együtt fejlődnek. Úgy véljük tehát, hogy az emlősállat igen sok oly cselekvésre képes, mely megtanulást, megértést, emlékezetbentartást követel, sok ily cselekvést kombinálni is tud; az elvont felfogás képessége azonban hiányzik, vagy igen csekély s így képtelen oly cselekvésekre, amelyek ezen a képességen alapszanak”. Vogt és Brodmann újabban oly kísérleteket végeztek, melyek arra engednek következtetni, hogy az agy falilebenyeinek melyek az embernél ugyancsak tekintélyes nagyságot érnek el, szintén lényeges jelentőségük van a magasabb lelki tevékenységek keletkezésében. Sőt, mi több: a két fentnevezett agykutató szabatos kutatásai arra az eredményre vezettek, hogy az agyközpontok sokoldalú kialakulása, a részleges szervek szöveteinek nagyító alatt megfigyelhető differenciálódása az, ami az embert megkülönbözteti az állattól. Az ember agyvelejében sokszorosan több különleges központ van, amelyek nem alacsonyrendű érzékelő vagy mozgási központok. Ami az állatoknál, még az emberszabású majmoknál is egyenletesen felépített agymező, az az embernél több, jól megkülönböztethető alközpontra oszlik. Az embernél magában, a homlokagyban eddigelé mintegy 70 ilyen központot mutattak ki. Az állatoknál, még az emberszabású majmoknál is számuk legfeljebb 12! A nagy agykéreg egész felületének az embernél legfeljebb 20 százaléka esik az alacsonyabbrendű, érzéki központok teljes felületére, míg 80 százalék föléjük rendelt, magasabb tevékenységeknek szolgál. Ha egy átlagmajmot veszünk az óvilági majmoknak abból a csoportjából, melyeknek farkuk van, a viszony éppen fordított; már ez is ily mélyen áll az ember alatt!
Heck így folytatja ezt a gondolatmenetet, amikor áttér az ember és állat közti főkülönbségre, a beszédre: „Az emberi és állati értelem közt határvonalat kell húznunk; fogalmakkal dolgozó, elvont gondolkodás az állatoknál is hiányzik és ezért a beszéd képessége is, mely a fogalmakkal rendelkező elvont gondolkodás legbiztosabb bizonyítéka. Az állatoknál is megtaláljuk a beszéd bizonyos elemeit, az emberi értelembe vett tulajdonképpeni beszéd legalacsonyabb fokát, amennyiben képesek arra, hogy kedélyhullámzásaikat hangokkal fejezzék ki; legjobb esetben csak bizonyos elképzeléseket tudnak kifejezni, amelyek érzéki észrevételekkel és ebből eredő kedélyhullámzásokkal függnek össze. A magasabb értelemben vett beszédnek logikusan tagolt szó- és mondatalakjai vannak és fogalmakkal dolgozik: ezt az állatok sohasem érik el, egyszerűen azért nem, mert fogalmakkal dolgozó gondolkozásuk sincs, már pedig a beszéd ennek kifejező eszköze. Wundt azt mondja: „Arra a kérdésre, hogy miért nem beszélnek az állatok, leghelyesebb az a közismert válasz: mert nincs mit mondaniok”. Ehhez még a következőket fűzhetjük: mindezek után már nem csodálkozhatunk azon, hogy a beszédben szereplő szervekben, mint a gége, nyelv, stb., semmiféle oly különleges berendezést sem találunk, amiből ezeknek a szerveknek különleges képességére következtetni lehetne. A tagolt beszéd nem az emberi gégének és nyelvnek működése, hanem az emberi agy munkájának eredménye. Csakugyan, agyunkban megtaláljuk a beszélőképesség központját, mégpedig a harmadik homloktekervényben; ez az úgynevezett Broca-féle központ. Ez azonban csak a tulajdonképpeni beszéd szolgálatában áll; ehhez járul egy másik központ a halántéklebenyben, valószínűleg az első halántéktekervényben; ezt Wernicke fedezte fel 1880-ban; feladata a megértés. Ezek után világos, hogy hogyan kell értelmeznünk azt a tényt, hogy az emberek beszélnek, az állatok nem. Nem az állati gégében, hanem az állati agyvelőben van a hiány, és éppen ez a legjobb bizonyítéka annak, hogy az ember és állat szellemi élete közt mélyreható különbség van”.
„Ezzel szemben a magasabb és legmagasabb rendű állatokat két más lelki tevékenység sorakoztatja az ember legközvetlenebb közelébe; első tekintetre tán nem is látszanak különös, kimagasló tevékenységeknek. Ezek az álom és a játék. Sokszor tapasztaljuk, hogy a kutya álmában morog, vinnyog, elnyomott ugatást hallat, miközben farkát csóválja, végtagjait rángatja. Ugyanezt a lónál is megfigyelték. Mindebből az következik, hogy az állatok is álmodnak, hogy alvás közben a központi idegrendszerben valamilyen inger következtében olyan változások állnak elő, mint ébrenlét alkalmával az érzéki észrevételekkel kapcsolatos kedélyhullámzások és testmozgások. Persze nehéz megmondani – valószínűen sohasem fogjuk biztosan megállapíthatni –, hogy az állati álmok mennyire közelítik meg az emberi álomban függetlenül működő fantáziát. Mindenesetre azonban nagy az állati álom jelentősége, amennyiben belülről kiinduló lelki tevékenységnek látszik, amelynek keletkezésében kimutatható külső ingerek nem vesznek részt”.
„Az álmoknál jobban ismerjük az állatok játékait és ezzel kapcsolatban is könnyű felismerni a különbséget az ember és az állatok közt. Elfogulatlan megfigyelések bizonyítják, hogy az állatok játéka a legelemibb életműködések játékos utánzásában merül ki: ilyenek a táplálékszerzés és életfenntartás; játékuk tehát vadászat, harc és menekülés. Ha lelki szemeink elé idézünk játszó állatokat, akkor fölösleges ezt az állítást bizonyítani. Jóllehet az állati játék tartalma ily korlátozott, mégis értékelnünk kell azt a jelenséget, hogy a magasabbrendű állat bizonyos kezdetleges képzelőtehetség segítségével komoly, vad ösztöneit, egész az ártatlan játékig le tudja gyöngíteni. Azon sem kell csodálkoznunk, hogy épp a háziállatok mutatják a legtöbb játékos kedvet, hisz természetes, vad ösztöneiket az emberi befolyás letompította.”
„A legújabb tudósítások és tapasztalatok megcáfolják, hogy az eszközök használata lényeges különbséget tenne ember és állat között. A magas szellemi színvonalon álló állat a fogságban, az ember példájának és befolyásának hatása alatt, megtanul eszközökkel bánni, föltéve, hogy testileg képes rá; ezt bizonyítják a megszelidített majmok és elefántok, végtagjaikkal, illetőleg orrmányukkal. A berlini állatkertben évekig élt egy japáni vörösarcú majom, mely – persze a neveletlen ifjúság példájára – pompásan tudott homokkal és kövekkel dobálózni és művészetét napról-napra gyakorolta a látogatók nagy örömére, mégpedig a legnagyobb dühvel és azzal a félre nem magyarázható szándékkal, hogy ellenfeleinek fájdalmat okozzon. Ugyanezt szavahihető utazók is tapasztalták, például az afrikai vad páviánok sziklák közt élő csordáiról, ugyanezt elmondja Neumann Oszkár is. Újabban Zenker, a kitűnő gyüjtő és gorillaismerő, Kamerunban figyelte meg, hogy az öreg gorillahímek letépett lombos ágakat használnak a legyek elkergetésére. Annyi tehát bizonyos, hogy az eszközök leleményes használatát ma már nem tekinthetjük oly szellemi képességnek, mely legkezdetlegesebb formájában is meg volna tagadva az állattól.”
„És végül – emlékezzünk meg – a lelki életnek arról a nagy világáról, amelyet kedélyhullámzások néven foglalunk össze! Csak általánosságban, a következőket szeretném megjegyezni: itt is megvannak ugyanazok az alapvonások, és alapelemek, mint az embernél; csakhogy minden kevésbé világos és tudatos, mert hiányzik a fogalmi gondolkozás. Mindazonáltal sok tekintetben igen magas és finom fejlettségi fok érhető el, és ezen nem szabad csodálkoznunk. Mert, ha a modern természettudományos felfogás hatalmas tényezőinek – ezek: az alkalmazkodás és a természetes kiválogatódás a létért való küzdelemben – a testi fejlődésben oly csodálatos alakító erőt tulajdonítunk, amely a növény- és állatvilág oly végtelen sokféle alakjában áll előttünk, akkor ugyanezeknek a tényezőknek alakító erejét el kell fogadnunk a lelki életre vonatkozóan is, és el kell ismernünk, hogy ezen az úton igen sok ösztönöset tenyésztett ki a természet, ami az egyes esetekben első pillanatra magasfejlettségű lelkiélet tudatos kifejezőjének látszik.”
„Ha elfogulatlanul és tudományos föltevésnélküliséggel akarunk eljárni, akkor az ösztönszerű kitenyésztődésnek ezzel a természettudományos magyarázatával igyekeznünk kell megérteni az állatok lelki életének legfinomabb és legnemesebb megnyilatkozásait is, amelyek másfelől éppen az erkölcs és erkölcsiség kezdeteinek látszanak. Én természettudományos szempontból – s ez az egyedüli helyes szempont itt, az állatok megítélésénél – az erkölcsöt és erkölcsiséget úgy fogom fel, mint a naiv, nyers önzésnek, az egyéni jólétre való kiméletlen törtetésnek elnyomását, elnyomását annak a természetes törekvésnek, mely az önfenntartási ösztönből kifolyóan minden szervezet sajátja, s ez a közösség javának érdekében történik. Természetesen ilyesmi csak társas állatoknál fejlődhetik ki, mert csak ezeknél találunk magasabb közösséget. Társas állatoknál – ha egyébként is szellemileg elég magas fokon állnak – meg is találjuk az erkölcsnek ez első nyomait, amennyiben az állat saját egyéni javát a köz java mögé állítja. Emlékezzünk csak a majomcsapatokra, az elefántnyájakra és belső, jól rendezett életükre, vagy a kutyára, amelynél – mint ahogy ebben az összefüggésben röviden mondhatjuk – saját fajának közössége helyébe a gazda lép, akinek magát alárendeli.”
Volt idő – s ez az idő belenyúlt a múlt század második felébe is –, mikor nagyon lebecsülték az állatok szellemi képességeit, és egyidejűleg – nevezetesen Németországban – az állatkedvelés és állattenyésztés is igen alacsony fokon állott. Azután, Darwin korszakalkotó művei nyomán, nagy változás és fellendülés következett be a tudományban, a művelt közönségnek legszélesebb köreiben pedig Brehm „Az állatok világa” című munkája nyomán. Nevezetesen Brehm, ez a zseniális író volt az, aki hangulatos is mélyérzésű leírásaival az állatot, mint élőlényt belopta minden olvasó szívébe. Őrizzük meg ezt a szeretetünket, de egyúttal tartsuk szem előtt a tudományos kritikát is és az előítéletektől ment, szigorúan tárgyilagos kutatást! Mert nem az a legjobb állatbarát és állatvédő, aki az állatot túlzó módon az ember mellé állítja és érzelgősen magasztalja, hanem az, aki becsületesen törekszik arra, hogy igazi állatismerő legyen, aki az állatot nem becsüli le, de túl sem értékeli.
Eddig megismertük az emlős szervezet legfontosabb és legjellemzőbb tulajdonságait. Vessünk egy pillantást az életmódra is, melyet az állat testi és szellemi képességei szabnak meg, szem előtt tartva azonban, hogy mindegyik állat a maga tehetségei szerint igazodik s a legteljesebb mértékben hasznát veszi természetadta képességeinek.
Az emlősök tulajdonképpen a szárazföld lakói, bár vannak köztük tengeri emlősök is, amelyek kizárólag vízben élnek. Ne felejtsük el azonban, hogy a tüdő – a tengeri emlősöké is – voltaképpen a szárazföldi élethez szükséges szerv. A viziélethez való alkalmazkodás néha annyira tökéletes, hogy külsőleg tökéletesen a halakhoz hasonlók, halformájúak, úgyannyira, hogy például a delfin, mely a legügyesebb halakkal is felveszi az úszásban a versenyt, a szárazföldön éppoly hamar elpusztul, mint a halak. A tengeri emlősök élete mégis elsősorban a levegőhöz és nem a vízhez kötött; bizonyos időközökben kénytelenek a víz felszínére szállani lélegzetvétel céljából, hogy az élethez nélkülözhetetlen oxigént a levegőből felvehessék. A víz csak állandó tartózkodási helyük a tengeri emlősöknek, amely megkönnyíti a nagytömegű test súlyát, s így hihetetlen nagy testsúllyal is, aránylag fürgén mozoghatnak. Ebből a szempontból tökéletesen mindegy, hogy víziemlőseink édes- vagy tengervízben élnek. A tenger óriásai, a bálnák, nemcsak az emlősök, hanem az állatvilág óriásai is, melyeket csak a tenger mérhetetlenül gazdag és kiapadhatatlan állatvilága tud a szükséges táplálékkal ellátni.
Ezzel ellentétben a legkisebbek azok az állatok, amelyek nagy erőfeszítéssel mozognak vagy szerzik táplálékukat; ilyenek a repülők és az ásók – a denevér és a vakondok –, vagy a rovarevők általában. Ezeknél a test tömege oly arányban kisebbedett, amilyenben a víziemlősöké megnövekedett.
Eltekintve az úszó, repülő és ásó emlősöktől, túlnyomó nagy részük a föld felszínén él. Az emlősök röghözkötöttsége sokkal feltűnőbb, mint a fürge, mozgékony madaraké, a lég igazi uraié; igen sok madár repülésközben dalol, ami ugyancsak azt bizonyítja, hogy a repülés igazi elemük. Ne felejtsük el azonban, hogy a madarakat a gyors emésztés és elválasztás következtében örökösen korgó gyomruk, és az előbbivel kapcsolatos magasabb testhőmérsékletük képesíti, sőt kényszeríti az állandó mozgásra, repülésre.
Az emlősök élete jóval nyugodtabb és kényelmesebb. Gyomrukat nyugodtan megtölthetik, s minthogy keveset mozognak, elegendő idejük van a nyugalomra és emésztésre. Evésből és pihenésből áll életük, melynek rendjét csak ellenségeik és néha az idő mostohasága zavarják meg, nem kellemes változatosságot vive életükbe.
Mindez azonban nem jelenti azt, hogy a mozgási képesség hiányzik az emlősállatokból. A legkülönfélébb mozgásoknak az emlősök között is vannak mesterei, művészei. A delfin éppoly jól úszik, mint a legügyesebb hal, a korai denevér ügyesség dolgában, ahogy a röpködő bogárságra vadászik, a fecskével versenyez; a vakondok éppoly gyorsan halad ásás közben a föld alatt, mintha a föld felszínén mozogna. A ló és az antilopok szélgyorsasággal száguldanak a steppék fölött és nagy távolságra nyargalásznak naponként, hogy szomjukat elolthassák; még az annyira nehézkesnek látszó elefánt is egy éjszaka alatt hihetetlen távolságot hagy maga mögött, és a kenguru, vagy akár a kicsiny ugró-egér is könnyű szerrel hagyja hátra üldözőjét, oly nagyokat és gyorsan tud ugrani menekülés közben. A ragadozók közül a zsákmányt meglapulva leső macskák hatalmas ugrással vetik magukat áldozatukra; a vad nyomában rohanó kutyát pedig hihetetlen kitartása jellemzi, amellyel előbb-utóbb sikerül legyőznie áldozatát. A legmagasabb fa hiába rejti el gyümölcsét, vagy az ágai közé rakott madárfészket a kúszni tudó emlősök elől; a gyümölcsöt kedvelő medve, a fészekrabló nyuszt és nyest, éppúgy, mint a mókus és egyéb rágcsálók kitünő kúszók, ugróművészek, sőt kötéltáncosok. A kúszásnak kiváló művészei a majmok is, a legmagasabb fákra éppoly ügyességgel másznak fel, mint a legmeredekebb sziklára. A sziklákon otthonosan mozog a kecske, a juh, a zerge pedig csodálatos biztonsággal és gyorsasággal száguld fel, a teljesen meredek és megmászhatatlannak látszó sziklafalon. Még a sarki tájak örök hó- és jéghonában is találunk emlősöket (jegesmedve, jegesróka, fókák, stb.), melyek csak ott érzik igazán otthon magukat.
Mint látjuk, az emlősök sincsenek híjával a mozgékonyságnak, mert járnak, ugranak, futnak, repülnek, kúsznak és úsznak, mindazonáltal jóval kevesebbet mozognak, mint a madár, amelynek a mozgás az igazi eleme.
Az emlősök tápláléka nagyon különböző, az állat- és növényvilág csaknem minden termékének akad kedvelője. A táplálék minőségével függ össze a fogazat alakja és igen gyakran a végtagok szerkezete. A táplálék minősége és a mozgás milyensége is szoros összefüggésben van egymással, együttesen nagy befolyást gyakorolnak az állat alakjára és a legcsodálatosabb alkalmazkodásra kényszerítik a szervezetet. Gondoljunk csak erszényes hangyász hosszú, csőalakú fejére és hosszú féregalakú nyelvére, az elefánt orrmányára, és az újvilági majmok kapaszkodó farkára; vagy a hegyi antilopok kemény és magas, a víziantilopok lágy és szélesen szétterpeszthető patáira; vagy a fakúszó borzok tapadós, a jegesmedve és a sarki nyúl erősen szőrös talpára. Mindannyi kitűnő példa az alkalmazkodásra.
A fogazat annyira jellemző egyes emlősökre, hogy az egyes csoportok elkülönítésében, vagyis a rendszertanban is igen nagy szerepet játszik. Így például a metsző fogak gyökértelensége és a szemfogak hiánya a rágcsálókat jellemzik; jól fejlett szemfoguk van a majmoknak és a ragadozóknak; viszont más zápfog alkalmas a húsevésre, más a rovarevésre és megint más a növényevésre.
A párzás ideje, mellyel a fiatalok gondozása áll viszonosságban, a legtöbb emlősnél meghatározott időhöz van kötve, mely szabályszerűen visszatérő változást hoz az emlős megszokott életébe. A fajfenntartó ösztön oly hatalmas, hogy még az önfenntartás ösztönét is elnyomja, úgyhogy az utódokról való gondoskodás – a legfontosabb feladat teljesítése – közben, a fajfenntartás ösztöne annyira úrrá lesz az állat fölött, hogy például a szarvas elfelejt legelni, az anyamedve pedig behavazott téli szállásán marad újszülött kicsinyei mellett hónapokon keresztül, a nélkül, hogy ez idő alatt enne valamit.
A madarak és emlősök között, páros életüket tekintve is, feltünő különbségeket találunk. Amíg a madarak általában egynejűségben élnek – e szabály alól csak a tyúkfélék egy része kivétel –, addig az emlősök majdnem mind a többnejűségnek hódolnak, a legtöbb hím a párzás után egyáltalán nem törődik többé a nősténnyel. Szorosabb, több mint egy évig tartó életközösségben valószínűen csak néhány kérődző, a kis antilóp fajok élnek.
A többnejűséggel kapcsolatos a másodlagos ivari bélyegek kialakulása, mely a hímet és a nőstényt már külsőleg is megkülönbözteti egymástól és amely különbség a többnejűségben élő madarakat is jellemzi; nevezetesen a hímek nagyobbak a nőstényeknél és különös fegyvereik is vannak. Ezekkel a fegyverekkel küzd a hím emlős a nászideje alatt a vetélytársakkal a nőstény birtokáért, mely küzdelemnek eredményeképpen a legerősebb nyeri el a pálmát.
A csoport vezetését, irányítását azonban nem mindig a legerősebb hím veszi át. A vadlovak között a vezérmén feltétlenül a legerősebb, a kérődzőknél azonban, különösen a szarvasféléknél, a vezetőállat, amely tényleg vezeti a csapatot és őrködik biztonság felett, mindig egy vén és tapasztalt nőstény; a szenvedélytől elvakult bőgőszarvas például nem is volna alkalmas a biztosító szolgálatra. A társas életet élő emlősök között a kötelességnek ilynemű elosztása valószínűleg azzal hozható összefüggésbe, hogy amíg a vezérmén állandóan a csapat mellett tartózkodik, addig a hím kérődzők csak a párzási idő tartamára csatlakoznak a csapathoz.
Érdekes, hogy a vén hímek nem mindig kelnek védelmére a ragadozóktól megtámadott csapatnak. Zell Tivadar figyelmeztet arra, hogy ez az állatok védelmi rendszerével függ össze, a szerint, hogy azok futással vagy támadással védekeznek a támadás ellen. Wissmann megfigyelése szerint a bika gondolkodás nélkül támad, hasonlóképpen – Brehm megfigyelése szerint – az öreg hím majom is.
A vemhesség ideje elég hosszú ideig tart, ezt követően a fiatalok táplálása és felnevelése is elég gondot ad, s így nem lehet csodálkozni, hogy a kölykök száma általában véve kevés. Egész rendekre (majmok, félmajmok, patások, tengeri emlősök) jellemző szabály, hogy rendesen csak egy fiat szülnek. Több mint 20–24 kölyköt (például a disznó) egy emlős sem szül egyszerre, bár a 8–10-es szám is elég gyakori. Az újszülöttek száma attól is függ, hogy milyen fejletten jön a kölyök a világra és mily módon szerzi meg anyja a táplálékot; a ragadozó nagyon fejletten születik, s már egy néhány óra mulva követheti anyját; az erszényes emlős jó ideig a hasán lévő bőrráncban – erszényében – hordja fiait.
Az emlősök hangja nagyon távol áll attól, mit kellemesnek lehet nevezni; csak a gibbon „énekel” félig-meddig „zenei” hanglétrán. A bőgőszarvas hangja, a vadat űző kutya csaholása csak a vadászfülnek kellemes; hasonlóképpen a türelmetlen paripa nyerítése is csak gazdájának tetszik igazán. Mindaddig azonban, míg különösebb jelentést nem magyarázunk a hangokra, olyan „dal” marad, „mely a követ is megindíthatja, s az embert őrültté teheti”. Gondoljunk csak a szamár ordítására, a kutya vonítására és a macska szerelmi nyávogására.
Általában az emlős ritkán hallatja hangját, a legtöbb esetben még a legnagyobb fájdalmat is hang nélkül tűri, sőt halál tusáját is némán vívja. A legtöbb hangot azonban mégis a fájdalom (nyúlsírás), az aggodalom és a félelem (őzböffenés, zergefütty) csalja ki belőlük, mely utóbbi figyelmeztetésül szolgál gondtalanul legelésző társaiknak is. A kutya ugatása ugyanebbe a csoportba tartozik, csakhogy az embert figyelmezteti a közelgő veszedelemre. A nász ideje alatt hallatott hívó hangok – szarvas bőgése – jóval erősebbek, ami könnyen érthető, ha az állat nagyfokú izgatottságát vesszük figyelembe; az őzbak hívására az őzsuta is megfelelő hanggal válaszol. A nőstény emlős – általában véve – külön hanggal csalogatja kicsinyeit, amellyel azoknak figyelmét akarja felkelteni; annak, aki szereti az állatokat, könnyen úgy hangzik az, mint a tudatos, büszke anyai szeretetnek megnyilvánulása.
A nőstény szülés előtt fokozott mértékben igyekszik lakást építeni, holott egyébként ez az ösztön nincs is benne nagyon kifejlődve. A lakásépítés a legtöbb esetben abból áll, hogy egy vacoknak alkalmas mélyedést keres az állat, melyet esetleg egy kis kaparással kényelmesebbé tágít. Amily nagy mester a madár, olyan kontár az emlős a fészeképítésben.
Csak néhány rágcsáló épít magának a föld fölött rendesebb lakást, ágakból és levelekből; a hód vára, a mókus nyári és téli lakása, valamint eleségkamrái, a törpe egér és a mogyorós pele gömbfészke, mintaszerű és a madárfészekkel teljesen egyenértékű mesterművek.
Egyébként az emlősök csak földalatti lakásokat építenek. A földalatti lakásépítők igazi mesterei a rágcsálók (marmota, üregi nyúl, hörcsög, egér, stb.). Vetélkednek velök a kúszni nem tudó, apróbb ragadozók is (sakál, róka, borz, mezei görény, stb.), a foghíjasok közül az övesállatok, a legszebb és legösszetettebb földalatti várat azonban állítólag a rovarevő vakondok építi.
A barlang- és fészeklakó-emlősöknek különös sajátossága, hogy a kedvezőtlen évszakokat – nálunk a telet – táplálkozás nélkül átalusszák; ezt a jelenséget, mely leginkább a költöző-madarak őszi vándorlásához hasonlítható, nevezzük téli álomnak. Télen csak a ragadozók, denevérek, rovarevők és rágcsálók rendjébe tartozó emlősök alszanak, ezek se mind, és téli álmuk sem egyforma. A medve téli álma alvás és bőjtölés; ezalatt minden testi tevékenység zavartalanul működik, az anyamedve magzatai szabályosan növekednek, a kicsinyek megszületnek és néhány heti, sőt hónapi szoptatásuk is „téli álomban” történik. A berlini állatkertben nem régen figyelték meg, hogy a terhes borz magzatának fejlődésében a téli álom alatt pihenési időszak áll be, vagyis a magzat növekedése szünetel. A mókus és a hörcsög téli álmából időnként felébred és éléskamráját felkeresve, táplálkozik is. A pele és marmota hideg halotti merevségbe esik; szívverésük meglassudik s ennek megfelelően lélekzetvételük is szembeszökően megcsappan, sőt félbe is szakad; a test melegsége alább száll, a végtagok megmerevednek és elhidegülnek; álmuk olyan, mint a tetszhalál, az életnek már csak nyoma látszik; végül a tetszhalott hirtelen felébresztése halált okoz.
Nagyon érdekesen igyekszenek a tél nyomorúságait elkerülni a denevérek; ha hiány áll be a repülő-rovarságban, egy részük téli álomra hajtja a fejét, mint a legtöbb emlős, más része vándorútra kel, mint a madár. Legújabban a denevérek vándorlását az Egyesült Államokban figyelték meg; e vándorlások teljes szabályszerűséggel következnek be az északi és déli államok fölött. Ez a megfigyelés ama régebbieket erősíti meg, melyeket Európában és Ázsiában is észleltek. De akadnak az emlősök között igazi vándorok is; ilyen az ó- és újvilági rénszarvas és ilyen volt – kipusztulása előtt – az amerikai bölény is. Majdnem évenkint vándorolnak a szibériai és északamerikai mókusok is; azonban ez esetben szabályos vándorlásról nem beszélhetünk, mert olyan helyen csődülnek tömegekbe az állatok, ahol elegendő a táplálék; állítólag egyes előfutárok kémkedik ki azt a vidéket, ahol a fák magvaiból jó termés van, s ahol azelőtt alig volt mókus, a mag érésekor halatlan tömegekben jelenik meg.
Így jutunk el az emlőstömegek egészen szabálytalan vándorlásaihoz, melyet a táplálék hiánya okoz. A külföldi irodalomban is híres, roppant tömegekbe verődő norvégiai lemming vándorlása is éhségre vezethető vissza és ma is elég gyakran látható. Történelmi emlék ma már a dél-afrikai antilopok vándorlása is; régi utazók napokon át látták vonulni egyik helyről a másikra mérhetetlen tömegeiket; az aranyásók és gyémántkeresők hamarosan véget vetettek ennek az idillikus állapotnak.
Nem vándorlás, hanem egyszerűen terjeszkedési jelenség a vándorpatkány elszaporodása Európában; az előnyomulás keletről-nyugatra, országról-országra, sőt helységről-helységre aktaszerűen bizonyítható.
Az emlős egész életmódja környezetéhez idomul. A környezet és állatkapcsolatát a színezet is felismerhetővé teszi. Kétségtelen, hogy az emlősöknél nem beszélhetünk oly pompás, díszes és ragyogó ruházatról, mint a madaraknál, de beszélhetünk olyan ruházatról, mely óvja, védi, elrejti az állatot ellenségei elől, még akkor is, ha az látszólag a legfeltűnőbb tarka ruhát viseli. A tigris és zebra keresztsávozottsága nagyszerűen beleillik a bambuszerdők nádtörzsei és a füves steppe levélformái közé; a leopárd és zsiráf foltjai a lombokon áthatoló napfény-foltokhoz hasonlítanak; minden utazó határozottan állítja, hogy ezeket a múzeumokban és állatkertekben oly tarkának és feltűnőnek látszó állatokat, kint a vadonban nagyon nehéz meglátni, és hogy már igen kis távolságban összeolvadnak környezetükkel. Minden vadász tudja, milyen nehéz a fekvő nyulat meglátni, bundájának színe annyira hasonló a talaj színéhez. Ha azonban az őznek rókavörös nyári ruhája a zöld mezőn messziről látható, arra azt mondhatjuk, hogy a vörös színt nem a megmívelt zöld mezőre szánta a természet, hanem inkább száraz, erdei, leveles területre. A színbeli alkalmazkodás ténye félreérthetetlen, a havas területeken élő, fehér bundájú állatoknál (jegesmedve, havasi kecske; a hermelin, jegesróka és havasi nyúl téli, fehér öltözete). A sivatagi állatok (a vadszamár, gazella, sivatagi rókák, ugróegerek, stb.) homokhoz hasonló színe is védőszínezet.
Az emlősök legtöbbje nappal van ébren és éjjel alszik, de azért csaknem minden rendben vannak nappali és éjjeli állatok. Az éjjeli emlősök között vannak valódi éjjeli állatok (félmajmok, rovarevők, erszényesek, stb.), melyeket bagolyszerű szemük is állandó éjjeli életre és nappali álomra utal; de vannak köztük olyanok is, amelyek a ragadozóktól való félelmükben járnak éjjel vagy estszürkületkor zsákmány után.
A kimondottan társaséletet élő emlősök mellett, remeteéletet élők is vannak. Egyes fajok csak a párzás és a fiatalok felnevelésének tartamára verődnek csapatba, és mihelyt elérték céljukat, a közösséget felbontják. Az időtartam, amely alatt az emlős-kölyök önállóságát elérheti, csaknem éppoly különböző, mint az állatok nagysága. A szárazföldi emlősök között, határozottan az embereknek kell a legtöbb idő, hogy kifejlődhessék; még az elefánt is előbb nő meg.
Az egyes emlőscsoportok az élet nehézségeinek leküzdésében, sokoldalúságuknál fogva, nagy mesterek; bár az emlősök állnak az állatvilág élén, azért sok tekintetben elmaradottak. Csak a teremtés koronája, az ember áll az emlősök felett, mert tudja mindazt, amit az emlősök tudnak; rosszabbul tud ugyan futni, mászni, kúszni, mint az emlősök, csak repülni nem tud, de mégis túlszárnyalja azokat szellemi képességeinek segítségével. Kérlelhetetlen szigorral uralkodik a földön és lakóin, ahol megjelenik, egy-kettőre megbontja a természet egyensúlyát, valóságos rémuralmat gyakorol, kultúrájának terjedését a gyengébb állatvilág pusztulása jelzi, s nem kis keserűséggel elmondhatjuk, hogy ma már csak ott élnek nagy emlősök, ahol az ember nem bírta vagy – ritkábban bár – valamilyen oknál fogva nem akarta kiírtani azokat. Minden természetbúvár világosan látja, hogy a kultúra terjedése az emlősöket sújtja a legjobban, és el fog jönni az az idő is, amikor az embert csak hasznos háziállatai fogják körülvenni a földön; az emberre közömbös, különösen pedig az ártatlan emlősök utolsó példányait legfeljebb kegyeletből fogja megtűrni.
Minél világosabban látjuk a jövőt, annál kevésbé látunk világosan visszafelé.
Az emlősök keletkezését és származását még mindig nem tudjuk biztosan; hogy az emlősöknek történetük és multjuk van, azt számtalan ásatag csontmaradvány bizonyítja. Legkönnyebben a jóval erősebb és ellenállóbb nagy emlősök csontjai kövesednek és maradnak meg, és az annyira fontos, legrégibb emlősöket, melyek kicsinyek, s így csontjaik törékenyebbek, csak egy néhány állkapocs, koponyatöredék és fog képviseli. Mert az emlősöknek legrégibb őseit és a hozzájuk vezető, átmeneti alakokat is, kicsinyeknek kell tartanunk; általánosan tapasztalt és közismert dolog, hogy minden állatcsoportban, a legrégibb, legegyszerűbb és legkezdetlegesebb formák egyszersmind a legkisebbek is. Az emlősök sem kivételek e szabály alól; a legrégibb emlős és emlőshöz hasonló maradványok, amelyeket eddig ismerünk, mind igen kicsiny állatok maradványai; így egy nyúlnagyságú állat koponyatöredéke és egy megkövesült lábnyom (Tritylodon, Theriodesmus), a felső Karroo-formációból, vagyis a délafrikai tábla középső teraszaiból származik; ezenkívül néhány apró zápfogat is ismerünk, a Dél-Német országban és Dél-Angliában egyformán meglevő, úgynevezett „bonebed” rétegből. Úgy a Karroo-formáció, mint a bonebed homokkőrétegek triászkorúak, vagyis földünk történetének második nagy fejezetéből származnak. Ily messze, vagy talán még messzebb kell keresnünk az emlősök eredetét, mert a legrégibb emlősök egyidejű előfordulása Németországban és Dél-Afrikában, azt bizonyítja, hogy azok a triász végén már nagyon el voltak terjedve a földön.

Dél-Afrikai Theriodonta-maradványok. Lycosaurus-koponya;
1. oldalról, 2. elülről, 3. egy fog külön.
Owen "Palaeontology" c. könyvéből.
Igen érdekes, hogy a legrégibb ásatag és a ma élő legalacsonyabbrendű – az ausztráliai csőrös – emlősök között szoros kapcsolat van, helyesebben néhány rendkívül fontos bélyegben egyeznek meg egymással. Tudjuk, hogy a csőrösemlősnek a rendkívül korán kihulló tejfogán kívül más – emlősfog értelemben vett – foga nincs. A kifejlett állat csőrén találunk ugyan szarufogakat, de ezeknek a rendes emlősfoghoz se alaktanilag, se fejlődéstanilag semmi köze sincs. A tejfog rendkívül apró, felülete homorú – tálszerűen bemélyedő –, melynek szélein apró, kis gumók sora foglal helyet. A csőrös emlős tejfoga rendkívül hasonlít a würtenbergi Echterdingen bonebed rétegeiben talált Microlestes fogához, továbbá az ugyanonnan előkerült Trigliphus, az angolországi Plagiaulax, a délafrikai Tritylodon és Theriodesmus fogaihoz, vagyis azokhoz a legrégibb emlősökhöz, melyeket Multituberculata (sokgumósfogúak) vagy Allotheria néven szokás, külön rendbe, összefoglalni. Az eddig ismert Multituberculata-maradványok alapján azt gondoljuk, hogy azok a ma élő csőrősemlősökhöz rendkívül hasonlók és valószínűen ugyancsak tojásrakók voltak.

A csőrösemlős más tulajdonságokban is eltér a rendes szabású emlősöktől; például szarunemű csőre van, amely tulajdonságok látszólag a madárhoz teszik hasonlóvá; azonban a valóságban a madarak és emlősök között semmiféle származástani összefüggés nincsen. A gerincesek között, a madarak és emlősök törzse, két egyforma értékű, de egymástól teljesen különböző, magasfejlettségű törzs, amelyek melegvérűségük által, a legmagasabb fejlettségi fokig jutottak.
A madarak közvetlen rokonságát a reptiliákkal (csúszómászókkal) már Huxley, Darwin kortársa, bebizonyította; Huxley vizsgálatait az Archeopterix felfedezése csak megerősítette, s egyelőre nem látszik valószínűnek, hogy a madarak származását illetően meg kell változtatnunk eddigi nézeteinket. Sokkal bizonytalanabb alapokon nyugszik Huxleynek az emlősök származásáról írt elmélete. Huxley ugyanis annak alapján, hogy úgy az emlősök, mint a kétéltűek koponyája két bütyökkel ízesül az első nyakcsigolyához, a két csoport között szorosabb származástani kapcsolatot tételezett fel. A modern törzs- és egyénfejlődéstani vizsgálatok azonban nem igazolták ezt a feltevést. Huxley elméletét Gaupp, a freiburgi egyetemen az anatómia tanára, vette bírálat alá és tanulmányainak eredményét „Az emlősök rokonsági viszonyai, különös tekintettel a koponya morfológiájára” címmel, az 1910-ben Grázban tartott zoológiai kongresszuson, adta elő. Gaupp nézete szerint a kettős nyakszirti bütyök, nem olyan alapvető különbség, hanem az egyszerű nyakszirti bütyökből fejlődött ki a két első nyakcsigolyának az emlősökre jellemző átalakulásával kapcsolatban. Más szóval az emlősöket is a reptiliákból kell származtatnunk és pedig olyan kihalt alakokból, amelyek a ma is élő reptiliák közül az újzeelandi Hatteria-hoz állnak legközelebb. Már régen rámutattak mások is, itt-ott, erre a lehetőségre. Így Flower az emlősök és reptiliák nyakcsigolyái között is talált bizonyos összefüggést, mert hiszen ismerünk két nyakszirti bütyökkel bíró ásatag reptiliákat – Theriodontia (emlősfogúak) – is, amelyeknek ráadásul fogazatuk is megközelítően épúgy metsző-, szem- és zápfogakra tagozódik, mint az emlősök fogazata. A fogak alakja, a munkabeosztás elvének megfelelően tehát először a Theriodontiá-nál különféle.
Annak, hogy az emlősök és a reptiliák között vannak átmeneti alakok, legszebb bizonyítéka az, hogy a Tritylodon nemet először a sokgumósfogú emlősök (Multituberculata) rendjébe sorolták; később az emlősfogú reptiliák rendjébe osztották be az állatot; Broom R. legújabb munkájában („Proc. Zool. Soc.” 1910) a Tritylodon típusának „leggondosabb tanulmányozása” után arra az eredményre jut, hogy az mégis a legrégibb – a sokgumósfogú – emlősök közé tartozik. Igen érdekes, hogy a Karroo-formációt – a Tritylodon anyakőzetét – Elő-Indiában is megtalálták, csakhogy ott Gondwana-formációnak nevezték el. A Karroo- és Gondwana-formáció tehát nem egyebek, mint egy rég tenger alá – és pedig az Indiai-óceán alá – süllyedt egykori világrész maradványai. Nem lehetetlen, hogy ezzel a földrésszel együtt, a tengerbe vesztek az emlősök származásának kétségtelen bizonyítékai is. Bármilyen is volt azonban az a bizonyos ős, kétségtelen, hogy emlőssé való átalakulását a bőr átalakulása tette lehetővé, amennyiben a test külső védelme mellett átvette a test hőmérsékletének szabályozását is. A hő megtartásának lehetőségével együtt járt a melegvérűség, amely nagyobb életenergiát, s így a továbbfejlődés lehetőségét jelentette. Jegyezzük még meg, hogy a csőrös emlősök és erszényesek testhőmérséklete jóval alacsonyabb, mint a többi emlősé.
Mai természettudományos felfogásunk szerint az emlősök eredetéhez és történetéhez szorosan kapcsolódik földrajzi elterjedésük. Mert az emlősök jelenlegi földrajzi elterjedése egyrészt a korábbi geológiai korokból átszármazott formák összességéből alakultak ki, másrészt a víz és a szárazföld elosztása és más fontos életkörülmények tették lehetővé, hogy a szó szoros értelmében „elterjedjenek” és pedig nemcsak egyszerű szaporodással, hanem kivándorlással is.
Az emlősök földrajzi elterjedésében az éghajlat és mindaz, ami vele összefügg – tehát az állatvilág tápláléka, a növényvilág is – fontos szerepet játszott, jóllehet, bizonyos az is, hogy megszokás és éghajlathoz való alkalmazkodás sohasem volt lehetetlen és gyakran meg is történt. Így például a földön majdnem mindenütt vannak rókák, s nagy elterjedésüknek megfelelően a legkülönbözőbb éghajlat alatt élnek. Általában véve az elmult földtörténeti korszakok mindezen viszonyainak megítélésében csakis a növény- és állatvilág nyujt támaszpontot, csak szárazföldi viszonyok megítélésében pedig éppen az emlősökre szokás következtetni, minthogy, főképpen a fiatalabb rétegekben, azok fordulnak elő a legnagyobb számban. Ebben az esetben tehát a zoológus és botanikus kevés hasznát veszi a geográfusnak, hiszen vizsgálataiban a geográfus is ő reájuk támaszkodik. Így tehát el kell fogadnunk azt az elméletet – amelyet éppen a növényi és állati maradványok ismerete alapján állítottak fel –, hogy földünk történetének ókora, pontosabban a kőkorszak óta egészen a mai napig a víz és szárazföld eloszlása nagyjában nem változott meg, vagyis az összefüggő szárazföld-tömegek már akkor is az északi féltekén voltak. Csak emlékeznünk kell arra, amit az emlősök eredetére vonatkozólag mondottunk, miszerint az emlősök első megjelenésének idejében, a föld ókora és középkora között, a triászban, Indiából még földhíd vezetett Afrikába, amely időközben az Indiai-óceánba merült.
A növény- és állatvilágnak – így az emlősöknek is – az északi nagy szárazföldeken lehetett az őshazája, oly területeken tehát, melyek minden ismert földkéregformáció kétharmadán át többé-kevésbé változatlanul maradtak fenn a jelenkorig. Régebben az volt a tudományos felfogás, hogy itt a létért való küzdelemben különösen megedzett, terjeszkedésre fölötte alkalmas formák alakultak, és ezek a sarkvidékről állandóan sugároztak, szivárogtak az egyenlítő felé; így beszéltek a szervezeteknek a sarkvidékről való meneküléséről, melyet egész természetesen a földkéreg lassú lehűlésével hoztak kapcsolatba. Ma azonban már kétségbe nem vonható, bár még meg nem magyarázott jégkorszak, ezt az egyszerű elképzelést lehetetlenné teszi, főkép akkor, ha ily lehűlési periódusokat nemcsak a harmadkor és jelenkor közt, hanem a föld ókorának (palaozoikum) végén is feltételezünk. Mai felfogásunk szerint az állandóan egymást követő elterjedési hullámok hajtották, szorították a növény- és állatformákat az északi sarkról az egyenlítő felé, sőt még azon túl is; és ezt nem külső okok idézték elő, hanem az a létért való küzdelemben kifejlett belső terjeszkedési erejükből kifolyóan történt. Ezt a belső erőt bizonyítja az a tapasztalati tény, hogy a szárazföldekről, kontinensekről származó alakok rövidesen elnyomják az elkülönült szigetformákat, ha velük találkoznak, amint azt behurcolt gyógynövényeknél, állati élősködőknél, a verébnél, a vándorpatkánynál és házinyúlnál gyakran megfigyelték. Nagy szárazföldek lakói közt ugyanis sokkal elkeseredettebb harc folyik a létért, mint valamely sziget sokkal kisebb számú lakói közt; azokat az állatokat és növényeket, melyek ily kemény iskolába jártak, az állandó küzdelem jobban megacélozta és erősebben fegyverezte fel új területek meghódítására.
Ha tehát a különböző földperiódusokban az állatokat és növényeket nem a földkéreg lassú lehűlésének külső kényszere hajtotta északról délre, hanem valamilyen belső kényszer – harcban edzett formáknak aktív törekvése további terjeszkedésre –, úgy ez a terjeszkedés nem állhatott meg a tropusokban, hanem még délebbre folytatódott, addig, amíg a földhidak vezettek. Csak oda nem juthattak tehát újabb és újabb vándorcsapatok, ahová földhidak – a tenger emelkedése vagy a földnek süllyedése következtében – nem vezettek.
Így egyszerre érthető, hogy a legfélreesőbb szárazföldeken és szigeteken, ha egyáltalán vannak emlősök, ezek – mint más jelek is bizonyítják – régibb eredetűek és a fejlődés alacsonyabb fokán állnak. A Csendes-óceán szigetein – ha csak Ausztráliához és Újguineához nem csatlakoznak – emlősök egyáltalán nincsenek, csak sertések és kutyák, melyek az emberrel kerültek oda. Hasonlóképpen az ősrégi Újzeeland kontinensen sincsenek emlősök, hacsak nem akarjuk elhinni azt a kósza hírt, amely szerint az ottani forró forrásoknál egy vidraszerű (tán helyesebben: csőrös emlős?) – a benszülöttek nyelvén a woitoteke-nek nevezett – állat él. Sclater, angol szisztematikus, aki először próbálta az állatföldrajz elemeit összefoglalni, ezeket a területeket ornithogaea-nak (madárföld) nevezte el, minthogy a fejlődés legmagasabb fokán álló lakói e területnek a madarak.
Az általános szabály alól mindenesetre kivételek a tengeri és légi emlősök, melyek vagy egészen függetlenek a szárazföldtől, mint a cetek és szirénák, vagy legalább is kevésbé függnek tőle, mint az úszólábúak és a denevérek, mert táplálékukat más közegben, a tengerben vagy a levegőben keresik és találják. A cetek elterjedését egyáltalán nem lehet kapcsolatba hozni az állatföldrajzi szárazföldbirodalmakkal és a szirénákról csak annyit mondhatunk, hogy az Atlanti- és Indiai-óceán tropikus partjain élnek. Már az úszólábúak és a denevérek inkább függnek a nagy szárazföldektől és szigeteiktől, s így földrajzi elterjedésük kötött, ámde közelről sem oly mértékben, mint ahogy azt a következőkben a többi emlősrendre vonatkozóan vázolni fogjuk. Kivételes helyzetük már abból is kitűnik, hogy füles fókák Újzeeland partjait is látogatják és denevérek nemcsak Újzeelandon, hanem a Csendes-óceán szigetein is vannak.
Az ornithogaea után a következő állatföldrajzi fokon Ausztrália és Újguinea áll, a hozzátartozó kisebb szigetekkel együtt, ahol az emlősök közül egereken kívül csak kloakás és erszényes állatokat találunk, tehát a legősibb emlősöket, azokat, melyek a fejlődés legalacsonyabb fokán állanak. Ezt a birodalmat, az ornithogaeával együtt notogaea-nak nevezzük.
Hogy az állatföldrajzi felosztást folytathassuk, meg kell ismerkednünk az átmeneti területek fogalmával. Az indo-ausztráliai vagy ausztromaláji átmeneti terület Újguineától nyugatra, a Szunda- és Molukka-szigeteken át bezáróan Celebeszig terjed, ahol még egy erszényes nem (genus) él. Wallace, az angol állatgeográfus az egymáshoz oly közel fekvő Borneo- és Celebes-szigetek közt határvonalat húz, mely élesen elválasztja egymástól az indiai és ausztráliai állatvilágot; Weber Miksa, az amsterdami emlős búvár a Wallace-féle vonal fogalmát újabban módosítja, és azt mondja, hogy a határon túl újabb átmeneti terület fekszik, amelyben, nyugatról keletre haladva mind kevesebb az ázsiai és mind több az ausztráliai állat.
Minthogy Dél-Amerikában is élnek erszényes állatok, azért mind általánosabbá válik az a felfogás, hogy Ausztrália valaha összefüggött azzal a Dél-Amerikával, mely neogaea néven újabb állatföldrajzi birodalmat alkot. Igaz azonban, hogy az újabban felfedezett Caenolestes kivételével csak erszényes patkányokat (Didelphiidae) találunk Dél-Amerikában, ezek pedig oly erszényes családhoz tartoznak, amely valaha Európában és Észak-Amerikában is élt. Mégis, mivel csak harmadsorban, tehát a földkéreg újkorában találjuk őket Európában és Észak-Amerikában, természetesnek találjuk, hogy ezek délről vándoroltak be Dél-Amerikába, ahonnan később az Egyesült-Államokba is eljutottak. Egyébként Dél-Amerika emlős világa igen gazdag, benne a félmajmok és rovarevők kivételével, minden rend képviselve van. Ha nem is egyezik meg az ó- és újvilág faunája, bizonyos hasonlóság, párhuzamosság mégis van közöttük, nevezetesen a majmok, ragadozók, rágcsálók, patások, különböző alakokban bár, de mindkét világrészben egyaránt előfordulnak. Vogt Károly szerint így van ez már a föld újkorának kezdete, az eocén óta, és ez csak úgy értelmezhető, hogy az emlősök a fentemlített északi őshazából egy északi összekötő szárazföldön át vándoroltak Amerikába s itt aztán önállóan és függetlenül fejlődtek tovább. Észak-Amerikát több ízben lepte el ily bevándorlási áradat, melyek közül az utolsók már nem érték el Dél-Amerikát; így magyarázható, hogy bár a Panama-szoros összeköti Amerika északi és déli részét, állatviláguk mégis lényegesen különböző. Az utolsó bevándorlás nyomai világosan kimutathatók, amennyiben a bevándorlók leszármazottainak legközelebbi rokonait Észak-Európában és Ázsiában is megtaláljuk; ilyenek a bölény, jávorszarvas, wapiti szarvas, havasi kecske, a rettentő vagy szürke medve (grizzly), baribál, róka és farkas, mormota és mások.
Míg Amerika állatvilága szerves kapcsolatban van Európa faunájával, addig az Antillák teljesen önálló állatföldrajzi területet alkotnak, éppen emlőseikre való tekintettel. Már Trinidad szigetén, mely az Orinoko területe előtt fekszik, s nincs messzebb a kontinenstől, mint Anglia Európától, feltűnő a különbség: a majmok, ragadozók, foghíjasok teljesen hiányzanak, ellenben az Antillákon (Kuba) – és csakis itt – egy sajátságos rovarevő nem (Solenodon) él. Hasonlókép a csodálatos nagy kúszópatkányok (Capromys) csakis az Antillákon fordulnak elő.
Észak-Amerika már beletartozik abba az óriási, összefüggő, már többször említett szárazföldtömegbe, mely az északi saroktól az egyenlítőig és azon túl terjed, s amelyet újabban sarki birodalom, arktogaea néven foglalnak össze. Itt találjuk a szárazföldi állatoknak, így az emlősöknek is főzömét. Minden rendet (ordo) megtalálunk itt, kivéve az erszényes- és csőrös emlősöket; ennélfogva a sarki birodalmat a magasabb rendű emlősök őshazájának kell tekintenünk. Természetes azonban, hogy ennek az óriási területnek különböző részeiben többé-kevésbbé mélyreható különbségek észlelhetők s ezért az arktogaeát, az emlősökre vonatkozóan is, négy tartományra osztjuk:
1. Holarktikus tartomány, melybe egyrészt Eurasia – azaz Európa és a vele éghajlatban, állat- és növényvilágban megegyező Észak- és Közép-Ázsia egészen Japánig – másrészt Észak-Amerika tartozik; ezek szerint a holartikus régió két altartományra: a palaearktikus (észak-óvilági) és nearktikus (észak-újvilági) subregióra oszlik. Minthogy ezt a nagy területet, egy tartományba foglaljuk össze, feleslegessé vált a régebben felvett cirkumpoláris (sarkkörüli) tartomány, melynek határa az északi sarkkör volna; a tartománynak két altartományra osztása teljesen jogosult, még ha az ó- és újvilágban uralkodó hasonlóságokat, párhuzamosságokat figyelembe is vesszük. Hisz igen sok család közösen előfordul mind a palaearktikus, mind a nearktikus altartományban, azonban más nemeket és fajokat alkotva. Így a rovarevők közül a cickány és vakondok; a ragadozók közül a valódi hiúz, farkas és róka, a medve, a tengeri vidra, a torkos rozsomák, borz, a tulajdonképpeni nyest és menyét; az úszólábúak közül a rozmár; a rágcsálók közül a pofazacskós mókus, mormota, ürge, a hód, a pocok, lemming, ugróegér, pocoknyúl és nyúl. A patások közül a bölény, kecskeantilóp, a vadjuh és szarvas különösen feltűnővé teszik, hogy a két altartomány egy felsőbb egységet alkot. Ezzel szemben a kecskék, minthogy csak az óvilágban élnek, jogosulttá teszik a két altartomány megkülönböztetését; hasonlókép a szarvasfélék közül a mosuszállatok csak Kelet-Ázsiában élnek, viszont a csodálatos villásszarvú antilopok, melyek a rendszerben egész különálló helyet foglalnak el, csak Észak-Amerika nyugati részén fordulnak elő. Tulajdonképpeni sertés az újvilágban nincs, helyettesíti azonban a pekari. A rágcsálók közül csak óvilágiak – palearktikusak – a pele-félék, a tulajdonképpeni hörcsögök, az ugróegerek és a lófejű egerek; a ragadozók közül a mosómedvék újvilágiak, a macskamedvék óvilágiak.
Mindkét altartományhoz egy-egy átmeneti tartomány csatlakozik, kevert állatvilággal: a nearktikushoz a szonóri átmeneti tartomány (nevét Szonorától, Mexikó legészaknyugatibb államától kapta), mely összekötő kapocs Észak- és Dél-Amerika között, tehát teljesen más állatföldrajzi tartományhoz, a neogaeához vezet; a palearktikus altartományhoz a mediterrán (földközi-tengeri) átmeneti tartomány csatlakozik, Európát Afrikával köti össze, vagyis az arktogaea két tartományát: a holoarktikusat az aethiopiaival.
A szonóri átmeneti tartomány, állatvilága alapján, már határozottan a délamerikai (neogaea) birodalomhoz tartozik és még sem szabad csak Észak-Amerika legdélibb részeit és Közép-Amerikát értenünk alatta, mert hiszen már az északi szélesség 43°-nál, New Yorktól északra kezdődik és Kanadába is benyúlik. A nearktikus altartományból így valóban kevés marad s ebből láthatjuk azt is, hogy az állatföldrajzban általában milyen nehéz tiszta fogalmakhoz jutni. Kétségtelen, hogy délamerikai alakok ily magas szélességi fokra is felhúzódnak; így például a puma, mosómedve, skunk stb. Másrészt azonban éppen emlőstörténeti fontos okok követelik, hogy Dél-Amerikát, mint önálló állatföldrajzi birodalmat (neogaea) tárgyaljuk.
Nagyon messze vezetne, ha a mai mediterrán átmeneti tartomány jelentőségéről a földtörténeti mult megvilágításában akarnánk valamelyes képet adni, vagy ha méltatni akarnánk azt a fontos szerepet, melyet a jelent megelőző földperiódusban, a harmadkorban éppen az emlősvilág kialakításában és elosztásában játszott. Csak megemlítjük, hogy a Földközi-tengert valaha földhidak szelték át s csak természetes, hogy ezeken a szárazföldi állatok átvándoroltak. A sok lelőhely közül csak kettőt említünk: harmadkori emlős maradványok tömegét találták Pikermiben, Athén és Marathon között és a Siwalik dombokban, a Himalaja déli lábánál; ez azt bizonyítja, hogy Afrika emlősvilágát, emberszabású majmait, antilopokban való gazdagságát és óriási zsiráfjait, elefántjait, orrszarvúit és vizilovait Európából és Ázsiából kapta; feltételezzük, hogy a meleg harmadkort követő jégkorszak, a hideg űzte ezeket a meleget szerető állatokat Európából és Indából a forró megkímélt Afrikába. Hogy az állatok ily nagy vándorlásokra is képesek, azt a Hemitragus vadkecskenem elterjedése bizonyítja, melynek két faja Indiában, egy faja Arábiában él.
Ma a mediterrán átmeneti tartomány – melyhez Dél-Európa, Afrikának a Szaharától északra eső része Afganisztánig és egész Elő-Ázsia is Beludzsisztánig tartozik – az állatvilágnak elég tarka képét mutatja. Európa, Afrika és Ázsia (India) – állatföldrajzi elnevezés szerint a holartikus, az aethiopiai és a keleti (orientális) tartomány – állatai találkoznak itt. A többi között itt él egy olyan majom is, mely Észak-Afrikából még Gibraltárra (Európa) is elterjedt (igaz, ott most már mesterségesen tartják fenn), a farkatlan magót, a keletázsiai, helyesebben indiai makakó-családból. Viszont és hasonlóképpen a gímszarvas legdélibb előfordulását az Atlasz vidékén (ma már csak a paratölgyerdőkben, Tunisz és Algir határán) találjuk, ahol ez az állat oroszlánok, leopárdok és csíkos hiénák között él. Az európai és az afrikai szarvasterületek között mindössze csak Szardiniában él egy törpe (szigetlakó) szarvasfaj. A sörényes juh – az egyedüli afrikai vadjuh, legközelebbi rokona a Szardiniában és Korzikában élő mufflon – Tuniszban és Tripoliszban a tehénantiloppal és mendeszantiloppal (Bubolis, Addax) találkozik, melyeknek rokonai, a többi antilopok Afrikában, a Szaharától délre élnek. Ezzel szemben az antilopok családjába tartozó gazellák tulajdonképpeni hazája a mediterrán tartomány sivatagjai és steppéi; a dámszarvas (másik ritka faj Perzsiában) hazája ugyanitt az erdőkben, a beozárkecske hazája pedig a hegységekben van. Más vadkecskék, zergék Spanyolországban és a Sinai félszigeten fordulnak elő; vadjuhok Észak-Afrikán kívül Szardinia és Korzika szigetén és egész Elő-Ázsiában is élnek egészen az Indus vidékéig. Afrikából egy ichneumonfaj egész Spanyolországig hatolt, sőt egy petymeg- (Genetta) faj Franciaországig is előnyomult; keleten a sárga, belsőázsiai vadszamár egész Turkesztánig és Transzkaspiáig hatol a mediterrán tartományba. Valóban gazdag emlősvilág: minden határból valami. Lydekker a mediterrán átmeneti tartományt önálló tartománynak szeretné venni; ezáltal sem válnék azonban egységesebbé.
2. Négerföldi (aethiopiai) tartomány, azaz Afrika a Szaharától délre. Ettől a tartománytól nem vitatható el bizonyos egységesség. Hisz ennek a tartománynak vannak a legtökéletesebb határai, mert a széles északafrikai sivatagöv olyan határ, akár az Indiai- vagy Atlanti-óceán és egy hasonló – az életre kedvezőtlen – sáv húzódik Arábián és Ázsián át is. Igaz ugyan, hogy a Szahara, kőzeteinek tanusága szerint, régtől fogva – részben a legrégibb kőszénkorszak óta – mint szárazföld emelkedik ki a tengerből, tehát ép oly régi szárazföld, mint Afrika többi része. Mégis pusztasága és növényekben való szegénysége miatt épp úgy akadályozta az állatvilág elterjedését, mint a tenger; különben hogy lehetne megérteni azt, hogy – az alakok egyformaságából és közeli rokonságából ítélve – Afrika állatvilágát Indiából kapta, mint ahogy azt az előbb már megemlített Siwalik-dombok gazdag harmadkori lelőhelye is bizonyítja.
A csimpánzok, páviánok, orrszarvúak és vízilovak, a zebrák, zsiráfok, bivalyok, antilopok, stb., mind Ázsiából vándoroltak be, s a mai Nílus-völgyén húzódtak délre. Itt tág terük volt a továbbfejlődésre, az új területen számos új faj keletkezett; így alakult ki az a fölötte gazdag, nagyszerű emlősvilág, melyet – a ráktérítőtől délre – Közép- és Dél-Afrikában annyira csodálunk, de amelyet „kultúránkkal” ugyanannyira veszélyeztetünk, sőt részben már ki is pusztítottunk. Valóban, Afrika ma hű képe egy jelenig megmaradt harmadkornak, vagyis olyan, mint amilyen hazánk is lehetett ebben az elmúlt földperiodusban, amikor még nálunk is jóval melegebben sütött a nap.
Éppen az emlősfauna gazdagsága a négerföldi tartományt legrövidebben azokkal az alakokkal jellemezhetjük, amelyek hiányoznak; ilyenek elsősorban a medvék és szarvasok. Nehezen érthető, hogy miért nem találták meg az Afrikába vezető útat. Igaz, hogy medvéket – az ajakos medvéket (Melursus) kivéve – Elő-Indiában sem találunk, de annál több ott a szarvas; könnyebb megérteni azt, hogy a vadkecskék és vadjuhok is hiányzanak – kivéve egy abessziniai zergét és egy délarábiai félkecskét – mert a kecskék és juhok a magas hegységek állatai, tehát nehéz elgondolni, hogy forró mélyföldeken hosszasabban vándoroltak volna.
Egyébként feltűnő bizonyos párhuzamosság, hasonlóság Indiával, az orientális régióval, ami várható is jól indokolt feltevésünk alapján. Afrikában csimpánzok és gorillák, Indiában az orangutang, hasonlókép kacskakezű majmok (Colobus) – karcsú majmok (Semnopithecus), makákók, páviánok – nagyszakállú makákók (Vetulus). Az ősi félmajmok közül a nyugatafrikai potto (Perodicticus) és medvemaki (Arctocebus) nemek élénken emlékeztetnek az indiai lajhármakira. A felsorolt adatok világosan szemléltetik az emlősállatok elterjedésére vonatkozó ama felfogásunkat, hogy a földtörténetileg mindig fiatalabb formáknak egymást követő elterjedési hullámai következtében, a legidősebb alakokat mindig a legtávolabbi területeken találjuk. Ugyanez a párhuzamosság feltűnően ismétlődik az indiai kancsilok (Meminna, Tragulus) és nyugatafrikai patkányok (Hyaemoschus) közt; jellemző, hogy ez utóbbit Dorcatherium néven előbb ismerték, mint ásatag állatot a rajnamenti és francia miocén (középharmadkor) korból és csak később találták meg, mint élő Hyaemoschus-t. Ez a – legtágabb értelemben – szarvasféle állatka Afrikában egészen egyedül áll, míg az előbb említett két félmajomhoz a Galago csatlakozik, mely egészen a keleti partokig előfordul. Ettől eltekintve, Nyugat-Afrikára fölötte jellemző a három fentemlített ősrégi emlős, mely az egyébként itt található állatvilággal nem igen áll összhangban; ezért jogosult, hogy Reichenow berlini ornitológus egy nyugatafrikai – a Guineai-öböl körül fekvő – erdőstartományról beszél.
Kizárólag Afrikában élő rovarevő állatok, az orrmányos cickányok (Macroscelides, Rhynchocyon), továbbá az aranyosszőrű vakondok (Chrysochloris), melyek közeli kapcsolatot mutatnak a Madagaszkár-szigetén elterjedt tanrek-kel (Centetes); Afrika sajátja még a Potamogale is, mely szorosabban szintén csak Madagaszkár Geogale nemével függ össze; a Potamogale csodálatos víziállat, oldalt összenyomott evezőfarokkal, mely zápfogainak háromcsúcsú (tritubercularis) alakjával a legrégibb emlősállatokra – a krétakori emlősökre – utal. Polamogale is csak a nyugatafrikai erdős tartományban fordul elő.
A ragadozók elterjedése is párhuzamos Afrika és India között. Így a cibetmacska-félék (Viverridae) közül a cibetmacskának (Viverra) és az ichneumon-nak (Herpestes) mind Afrikában, mind Indiában élnek megfelelő fajai, a petymegek (Genetta) ellenben kizárólag Afrika állatai. Csak egy, a többitől elütő alak, a Poiana él a Guineai-öbölben, Fernando Po szigetén, melynek legközelebbi, csak nehezen megkülönböztethető rokona a tigrispetymeg (Prionodon) Hátsóindiában található. Ez az eset is éles világítást vet azokra az állatföldrajzi vonatkozásokra, melyek a nyugatafrikai erdőöv és a keleti tartomány – nevezetesen annak maláji altartománya között – fennállnak. Másrészt Nyugat-Afrikában, de keletre egészen a Nyassza-földig található a tisztára indiai pálmasodróknak (Paradoxurus) egy már külsőleg is erősen különböző tagja, a nandinia, melyet testének felépítése alapján sok tekintetben a legprimitívebb, a fejlődés legalacsonyabb fokán álló ragadozónak tartanak. Végül a hiénaszerű ragadozók közül Afrikát kettő jellemzi és pedig a cibethiéna (Proteles) – mely a csíkoshiéna és cibetmacska között áll – és a tarka hiénakutya, egy hiénakülsejű vadkutyaféle, mely falkákban vadászik a nagy patásállatokra.
A valódi repülőmókusokat Afrikában a hozzájuk közel álló pikkelyesfarkú mókusok (Anomaluridae) helyettesítik, melyeknek farktövén két sor szarúpikkely van. Valódi hörcsögök nincsenek Afrikában, de ott él a hörcsögök családjához igen közel álló sörényes patkány (Lophiomys). A kizárólag délafrikai nyúlegeret (Pedetes) külseje után az ugróegerek (Dipodidae) családjába tartozónak nézhetnék, holott az utóbbiak csak Afrika mediterrán vidékein élnek; fogazatuk és más bélyegük alapján a pikkelyesfarkú mókusok mellé sorolják őket, de van olyan rendszertani író is, aki a sülalakúak csoportjába (Hystricomorpha) osztja be. A bojtosfarkú sülök (Atheura) jó példái annak, hogy a nyugatafrikai fajok alig különböznek a nekik megfelelő hátsóindiai fajoktól. A csalipatkányok (Octodontidae) családjánál a hódpatkányhoz hasonló nádi patkány (Thryonomys) tisztára aethiopiai, összes rokona azonban Dél-Amerikában él. Ilyen rokona a hódpatkány, melyet a szűcsök nutria néven ismernek. A csalipatkányok ilynemű elterjedése arra a gondolatra vezettek, hogy Dél-Afrikát és Dél-Amerikát valaha egy délsarki szárazföld – az Antarktika – kötötte össze egymással. A föltevés azonban, legalább a rágcsálókat tekintve, fölösleges, hiszen az Arktogaeában ásatag állapotban találták őket, tehát valószínű, hogy onnan származtak tovább.
Ugyanezt elmondhatjuk a foghíjasoknál is, melyek Weber szerint is három önálló rendre oszlanak; ezek közül tisztára négerföldiek a földi malacok (Tubulidentata) és négerföldi és orientálisok a pikkelyes állatok (Pholidota); ez utóbbiaknak megfelelő párhuzamos alakjaik élnek Afrikában és Indiában. Az afrikai elefántnak is megvan az indiai megfelelője. A páratlan ujjú patások közül az orrszarvú és a lovak között találunk párhuzamba állítható fajokat; így Afrikában zebrák és szürke vadszamarak, Indiában sárga vadszamarak és valódi őslovak élnek. A párosujjú patások közül a négerföldi és a keleti tartomány sertései és szarvasmarhái között már mélyreható különbségek vannak. Az antilopok nagy és alakokban oly gazdag csoportjában párhuzamosságról szó sincs: Indiában csak kevés képviselőjük van, Afrikában azonban az alakoknak oly gazdagságával találkozunk, mely, mint már említettük, a harmadkor patásállatokban való gazdagságára emlékeztet. A víziló ma kizárólag négerföldi, a történelmi időkben azonban még a Nílus alsó folyásának vidékén élt és ásatagon még messzebb is előfordul. Ugyanígy vagyunk a zsiráffal és ásatag rokonaival is, melyekhez még Afrika legújabb szenzációja, az ásatag világ élő képviselője, az okapi, csatlakozik. A kicsiny és rejtélyes Hyracoidea rend mormotaszerű külsejével és orrszarvúszerű csontrendszerével négerföldi, csak egyetlen faja terjedt el Szíriáig.
3. A madagaszkári tartományba csakis Madagaszkár szigete és a hozzátartozó kisebb Mauritius, Réunion, Rodriguez, Sechelles és Comora szigetek tartoznak. Mégis szükséges, hogy ezt a kis szigettartományt teljesen önállónak és az óvilág nagy állattartományaival egyenlő értékűnek tekintsük, mert egész állatvilága, különösen ami az emlősöket illeti, teljesen sajátságos. A szomszédos Afrika gazdag patásvilágából csak egy folyami disznót találunk itt; a víziló újra kihalt. Bizonyos, hogy mind a folyami disznó, mind a víziló átúszott ide Afrikából, amikor a kontinens és Madagaszkár közt elterülő, ma 1800 méter mély tenger még csak keskeny, sekély tengerszoros volt; ez a harmadkor vége felé, a pliocénben, vagy a pleistocénben történhetett. A négerföldnek minden későbbi, vagy a vizet kerülő és rosszabbul úszó bevándorlottja ettől a területtől el volt vágva. Régibb emlősök ellenben, így elsősorban a félmajmok, Madagaszkár szigetén elhatárolt területre találtak, ahol mai napig egyedül uralkodnak és ahol a nemeknek és fajoknak éppoly gazdagságára tehettek szert, mint a patásállatok Afrikában. Madagaszkár a félmajmok hazája, úgy, miként Ausztrália az erszényes állatoké; máshol csak elszórtan fordulnak elő. Madagaszkár szigetén a félmajmoknak több faja él, mint a többi emlősrendeknek együttvéve. A ragadozók közül csak cibetmacska-félék (Viverridae) élnek itt, melyek ősi, a fejlődés alacsony fokán álló alakok. Egyik nemüket (Eupleres) kezdetben rovarevőnek tartották, és az egyetlen itt élő nagyobb ragadozó, a fossza (Cryptoprocta) átmeneti alak a macskához macska-koponyával és macska-fogazattal, de cibetmacska lábakkal. Hasonlókép a sörtéssünök (Centetes) primitív sünök, melyeknek fogazata az erszényesekre utal. A sziget rágcsálói is, hörcsögszerű egerek, kizárólag Madagaszkár állatai.
Madagaszkár kicsiny, de önálló szárazföld, akárcsak Újzéland, ámde ennél sokkal fiatalabb; ilyen szempontból inkább az Antillákhoz hasonló. A rovarevők régi emlősrendjében a madagaszkári sörtés-sünök legközelebbi rokonai az Antillákban élő patkányvakondok-félék.
4. Indiai vagy orientális (keleti) tartomány. Minthogy a négerföldi tartomány emlősvilága sok tekintetben képmása és leszármazottja az indiai tartomány emlős faunájának, és erről az előbbivel kapcsolatban oly sok szó esett, a lehető legszűkebbre szabhatjuk mondani valónkat. Az indiai tartomány azonban nemcsak a négerföldivel áll szoros kapcsolatban, hanem széles területen érintkezik a holartikus tartománnyal is, ennek déli határa – a Himalája-hegység – révén. De összeköttetésben van az ausztrália-maláji átmeneti birodalom szétdarabolt szárazföldjének maradványaival, a földsüllyedés és beszakadások útján keletkezett szigetekkel is. Határai tehát elmosódtak és ezért – jóllehet állatvilága gazdag, – igen kevés olyan emlősállata van, mely kizárólagos sajátja. Ilyen ugyanis csak három van: a repülő maki, melyet mint külön rendet is (Dermoptera) szokás tárgyalni és ez esetben a denevérek és rovarevők között foglalnak helyet; a megnyult arcorrú, egyébként azonban mókus külsejű rovarevők családja a mókuscickányfélék (Tupajidae); végül a sajátságos kétkarmú félmajmok (Tarsiidae); mindkét utóbbi család egész kicsiny csoport, egy-két nemmel vagy fajjal. Mind a három csoport alakjai igen elütő, régi alakok, melyek főképpen a hátsóindiai-maláji altartományban élnek és amelyek az általános állatföldrajzi feltevésünknek – elméletünknek – megfelelően, az északi hazából beáramló elterjedési hullámok legtávolabbi végéhez tartoznak. Még fokozottabb mértékben érvényes ez a tapíroknak egymástól messze elszórt elterjedésére, melyek egyrészt az új birodalomhoz (neogaea) tartozó Dél- és Közép-Amerikában, másrészt az indiai birodalom hátsóindiai-maláji altartományában élnek. Általában, az orientális tartományra jellemző egyéb nemeket is mindinkább itt, mint az előindiai altartományban találjuk. Ilyenek: az orang-utan, a gibbonok, a nagyorrú majmok, a karcsú majmok, a lajhármaki, a karcsú lóri, a legtöbb előforduló kis ragadozó, a szarvasfélék közül a muntyákszarvasok és kancsilok. A medvék és szarvasok, melyek az aethiopiai tartománnyal ellentétben a keleti tartományra jellemzők, nagyjában egyenletesen oszlanak el az elő- és hátsóindiai altartományban; mind a két emlőscsoport a Himaláján át és északkelet felé összeköti az orientális tartományt a holarktikussal. Hasonlókép a félkecskék délarábiai fajukkal összekötik az orientális tartományt az aethiopiaival. A párosujjú patások közül a sertések és szarvasmarhák – a babirussza és a celbeszi ánoa eltérő nemeit kivéve – formákban igen gazdagok. Az antilopok ellenben messze hátramaradnak Afrika alakbősége mögött, viszont itt találjuk a csodálatos négyszarvú antilópokat.
Miután nagyjában megrajzoltuk az emlősök elterjedésének külső határvonalait, rajzoljuk meg az állatföldrajzi megfigyelések számára legkisebb egységnek, a sokat vitatott speciesnek vagy fajnak körvonalát. Úgylátszik, hogy az emlőstanban is számíthatunk ennek a rendszertani egységnek egész határozott földrajzi tartalmával; legalább is ezt vallja Matschie Pál, a berlini múzeum emlősgyüjteményének néhai kezelője. Matschie szerint az emlősállat függvénye szűkebb hazájának és ezen azt érti, hogy az emlősállat legfinomabb kialakulásában, hogy úgy mondjam, utolsó átformálódásában függ a környezetétől, külső életkörülményeitől, melyek a földfelület minden természetesen elhatárolt részein mások. Ebből következik, hogy Matschie a vízválasztóknak és az általuk elválasztott önálló folyamrendszereknek a fajképződésben alapvető fontosságot tulajdonít.
Ezek a kutatások még csak a kezdet kezdetén állnak és a dolog természetében rejlik, hogy tulajdonképpen csak akkor szabadna végérvényes ítéletet mondani, amikor már az egész emlősvilágot ebből a szempontból átdolgozták. Annyit azonban már ma is mondhatunk: azoknak az időknek örökre vége, amikor valamely emlősbőrre azt írták „Délafrika”, vagy éppen „Afrika”. Hiszen a modern rendszertan 1878 óta a megkülönböztethető emlősfajok számát 2000-ről 7000-re emelte! Ehhez csatlakozik még 4500 ásatag faj is. Nem tévedünk, ha feltesszük, hogy a legkisebb rendszertani-földrajzi egységre, a fajokra való hasadás, bár ugyanannak a törvénynek is van alávetve, a különböző emlősfajoknál különbözően – gyengébben vagy erősebben, elmosódottan vagy világosan – domborodik ki, a szerint, hogy ezek a formák mennyire hajlanak a változásokra. Így például mondhatjuk, hogy a tehénantilopok sokkal könnyebben megkülönböztethetők egymástól, mint a zöld cerkófmajmok és a zöld páviánok.
Azt is el kell fogadnunk, hogy az emlősfajok keletkezésében különböző okok játszanak közre. Ha egy magashegységben több főfolyónak forrása egymáshoz közel fekszik, akkor az ott élő magashegységi emlősök egymástól alig fognak különbözni, ellenben nagy különbséget találhatunk alföldekkel elválasztott magashegység-tömbökben, még ha egy és ugyanazon folyam vízrendszeréhez tartoznak is. Neumann Oszkár újabban ismét rámutatott arra, hogy ahol Abesszinia déli részén, a Hauasnak és a Kék-Nílusnak forrásvidékén mind a két folyó mellékfolyói keletről nyugatra egymáshoz közelednek, sőt egyesek össze is kötik a két folyót egymással, ott a Nílus és Hauas vízrendszerében bizonyára ugyanazokat az emlősformákat találjuk. Másrészt nagy folyamok elterjedési határokat alkotnak kicsiny emlősök számára, például a déloroszországi Dnyeper elválasztja a közönséges ürgét a gyöngyösürgétől, a Duna a magyar földi kutyát a szerémségitől. Ahol az idők folyamán a föld képe megváltozott, ahol folyók megváltoztatták medrüket, áttörések több folyó vízrendszerét egyesítették (Kongó, Zambézi), másokat pedig utólag elválasztottak, ott természetesen, az emlősök elterjedésének és fajképződésének viszonyai elmosódottak és bonyolultak. Így van ez Németország nyugati folyamrendszereinél (Rajna, Weser, Elba) és ez az oka, hogy a német emlősvilág nem adja oly világos példáját a vízválasztó-szabálynak, mint az afrikai emlősfauna.
Annyi azonban bizonyos, hogy a közgyüjtemények sem hasznavehetetlen lomtárak, mint sokan gondolják, és a sokat támadott rendszertani munka nemcsak a vizsgálati módszerek folytonos tökéletesedését vonja maga után, hanem ismereteinket is gyarapítja és pedig oly módon, hogy abból mélyebb megismerés is fakadhat; végeredményben pedig ez minden tudomány végső célja.
Végezetül az áttekintés megkönnyítésére, álljon itt az emlősök osztályának általunk követett rendszere; ugyanitt adjuk az egyes alosztályok rövid jellemzését is, melyet később nem ismétlünk; a rendszer a következő:
I. alosztály: EGYHÜVELYŰEK (MONODELPHIA)
Erszénycsontjaik nincsenek; az alsó állkapocs szögletnyulványa (processus angularis) – ha van – nem befelé hajlott; a két agyféltekét a kérges test (corpus callosum) köti össze egymással; hüvelyük egyszerű, a kettősség legfeljebb nyomokban jelentkezik; magzatuk méhlepénnyel fejlődik, emlőjük van; erszényük nincs.
ÖREGREND: FŐEMLŐSÖK (PRIMATES) (1. és 2. REND)
2. rend: Félmajmok (Prosimiae)
ÖREGREND: PATÁS ÁLLATOK (UNGULATA) (3.-TÓL 7. RENDIG)
3. rend: Párosujjú patás állatok (Artiodactyla)
4. rend: Páratlanujjú patás állatok (Perissodactyla)
5. rend: Talponjáró emlősök(Hyracoidea)
7. rend: Ormányos emlősök (Proboscidea)
9. rend: Ragadozó emlősök (Carnivora)
10. rend: Úszólábú emlősök (Pinnipedia)
11. rend: Rágcsáló emlősök (Rodentia)
12. rend: Újvilági foghíjas emlősök (Xenarthra)
13. rend: Pikkelyes vagy óvilági foghíjas emlősök (Pholidota)
14. rend: Csövesfogú emlősök (Tubulidentata)
15. rend: Denevérek (Chiroptera)
16. rend: Rovarevő emlősök (Insectivora)
II. alosztály: KÉTHÜVELYŰEK (DIDELPHIA)
Erszénycsontjaik vannak; az alsó állkapocs szögletnyujtványa (Proc. angularis) jellemzően befelé hajlott; az agyféltekék között kérges test nincs; hüvelyük kettős; a magzat rendesen méhlepény nélkül fejlődik, de kivételesen méhlepény is található; emlőjük, erszényük van.
17. rend: Erszényes emlősök (Marsupialia)
III. alosztály: MADÁRHÜVELYŰEK (ORNITHODELPHIA)
Erszénycsontjuk van; külön hollóorrcsontjuk van; lapockájuk síma, a tüske hiányzik; külön hüvelyük nincs; az emésztés, kiválasztás és ivarszervek közös kivezetője a kloaka; tojásrakó emlősök; emlőjük nincs, a tejmirigyek egyenként szájadzanak az úgynevezett szoptató mezőre.
18. rend: Kloakás emlősök (Monotremata)
A rendszertan ezt a csoportot Linné óta ismeri, aki az emberen, a majmokon és a félmajmokon kívül a denevéreket is a főemlősökhöz sorozta. Ismereteink gyarapodtával azonban mindinkább kiderült, hogy ez a beosztás nem helytálló, mert a félmajmok az igazi majmokkal semmiképp sem állanak valami közeli rokonságban, a denevérek pedig még kevésbbé. Ennek következtében nagyon is eltérő szervezetük alapján a denevéreket külön rendbe helyezték s a főemlősök csoportja szűkebb határok közé szorult: némelyek csak a majmokat sorozzák ma már ide, vagy a majmok közül is csak az emberszabású majmokat, mert hiszen a „főemlős” megjelölés, legalább bizonyos értelemben csak rájuk vonatkozhatik. Ma természetesen – miután felismertük, hogy az összes majmok magas fejlettségű agyvelejük mellett, teljes fogazatukban és ötujjú végtagjaikban nagyon is ősi vonásokat tüntetnek föl – már nem olyan könnyű róluk mint legmagasabb fejlettségű állatokról beszélni. Mi Webert fogjuk követni, aki a Primatesek csoportjában egyesíti a félmajmokat és a majmokat, s ezzel ennek a csoportnak olyan rangot biztosít a rendszerben, mint amelyet például a „patásállatok” elfoglalnak.
| 6. Változékonyság | TARTALOM | I. alosztály: Egyhüvelyűek (Monodelphia) |